Comparative Analysis of Pollen Morphological Traits in Halophytic Chenopods
-
Wafaa Kamal Taia

Department of Botany and Microbiology, Faculty of Science, Alexandria University, Alexandria, Egypt
| Received 23 May, 2025 |
Accepted 01 Jul, 2025 |
Published 10 Jul, 2025 |
Background and Objective: Halophytic plants exhibit various adaptive mechanisms to survive in high-salinity environments, one of which involves modifications in pollen morphology and internal tissues. However, limited information exists on the pollen traits of halophytes in saline habitats. This study aims to investigate the pollen morphological characteristics of halophytic species within the family Chenopodiaceae to understand their potential adaptive significance and evolutionary relationships. Materials and Methods: Pollen grains of 16 halophytic species belonging to 10 genera of Chenopodiaceae were collected from salt marsh habitats along the Western Coastal Region of Egypt, spanning from Alexandria to Mersa Matrouh, between October, 2018 and April, 2020. Pollen samples were prepared using acetolysis for light microscope examination; non-acetolysed pollen was sputter-coated with gold for scanning electron microscope investigation. Morphometric data, including pollen diameter and number of pores per pollen, were recorded and analyzed descriptively. Results: The study revealed notable variation in pollen diameter and number of pores per pollen grain, ranging from 12 to 62 pores. All studied species shared common features: Symmetric, apolar, spheroidal, zonopantoporate pollen grains with ornamented pores and a tectate imperforate exine. The exine was generally thin to moderately thick, with granulate or echinate surface textures. Based on pollen diameter, species were grouped into three categories: (1) Small grains 20 μm (e.g., Salsola longifolia, S. tetragona, S. volkanesii, (2) Medium grains 20-30 μm (e.g., Atriplex halimus, Anabasis articulate, Chenopodium album, and Chenopodium murale, and (3) Large grains 30 μm, including obligate halophytes and Basia muricata. Each group was further subdivided according to pore number: <20, 20-40, and>50 pores per pollen grain. Conclusion: The presence of small, pantoporate pollen grains with thin exine layers among these halophytic Chenopodiaceae species suggests a common evolutionary origin and possible adaptive modifications to salinity stress. These morphological features may contribute to their reproductive success in a saline environment and offer insights into phylogenetic relationships within the group.
| Copyright © 2025 Wafaa Kamal Taia. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Halophytic plants are those that can survive and adapt to hypersalinity. These plants have several ways to live under severe salinity. These plants have an economic importance in many ways, such as food fodders, chemicals, besides many others1,2. Genera belonging to the major angiosperm families containing the maximum number of halophytic taxa are the Chenopodiaceae s.l. (34 taxa; 22.08%), Poaceae (21 taxa; 13.64%), Fabaceae (14 taxa; 9.09%), and Asteraceae (13 taxa; 8.44%)3. In Egypt, Wanas et al.4 surveyed the halophytic taxa in the Damietta coastal area and found that members of Asteraceae precede those of the Chenopodiaceae, followed by the monocot taxa. Anyhow, these four families represent 53.25% of thehalophyteplants and have cosmopolitan distribution, and comprise of herbs or shrubs, rarely small trees or lianas. Among the worldwide distributed halophytic families is the Chenopodiaceae s.l. This is mostly cosmopolitan and tolerant with disturbed, arid to semiarid and/or saline habitats. They form an important component of the flora and vegetation of deserts and disturbed environments all over the world. The taxonomy of plants belonging to the family Chenopodiaceae s.l. (both Amaranthaceae s.s. and Chenopodiaceae s.s.) With over 170 genera and about 2050 species5 was considered as one of the core Centrospermous families, and has always been beyond dispute. The family is divided among ten subfamilies according to Hernández-Ledesma et al.6 into: Amaranthoideae, Betoideae, Camphorosmoideae, Chenopodioideae, Corispermoideae, Gomphrenoideae, Polycnemoideae, Salicornioideae, Salsoloideae, and Suaedoideae; and 15 tribes. But the most acceptable division of family Chenopodiaceae s.s. as mentioned by Kühn et al.7 is divided into four subfamilies with eleven tribes, viz., (1) Chenopodioideae with 6 tribes: Atripliceae, Beteae, Camphorosmeae, Chenopodieae, Corispermeae, Sclerolaeneae, (2) Polycnemoideae: with a single tribe Polycnemeae, (3) Salicornioideae: With 2 tribes: Halopeplideae, Salicorneieae, (4) Salsoloideae: With 2 tribes Salsoleae and Suaedeae. While family Amaranthaceae s.s. is divided into two subfamilies and four tribe8, (1) Amaranthoideae with two tribes: Amarantheae and Celosieae, (2) Gomphrinoideae with two tribes: Bryulineae and Gomphreneae. A great number of halophytes and xerohalophytes of the world belong to the Chenopodiaceae s.s.9-12. Genera belonging to this family have a fleshy nature and a pinkish color of the stem and sometimes leaves. According to the ecological classification13 the halophytes are under three categories; obligate halophytes, facultative halophytes and habitat indifferent halophytes. The obligate halophytic genera in this family are Arthrocnemon, Halocnemon, Kochia, and Sueada. Meanwhile, from the facultative halophytes the genera Chenopodium and Atriplex, while the genus Salsola considered from the habitat-indifferent halophytes. The genera Arthrocnemum, Atriplex, Bassia, Chenopodium, Halocnemum, Salsola, Salicornia, and Sueada are among the most common halophytic plants in the coastal regions of Egypt14.
Several studies have been done to investigate the pollen characters of the halophytes as those15-19. All the previous studies besides others, were mainly descriptive of the external features of the pollen taxa or for their identification. Differences in the pollen characters and types might provide a way to improve the taxonomic resolution within members of the Chenopodiaceae. Accordingly, selected common species from the previously mentioned genera, Arthrocnemum, Atriplex, Bassia, Chenopodium, Halocnemum, Salsola, Salicornia, and Sueada; have been subjected in this study to investigate the pollen morphological characters between the most common halophytic taxa in the coastal region of the Mediterranean Region in Egypt, from Alexandria to Mersa Matrouh Cities. Also, to understand if pollen characters between the halophytic categories can help in understanding their phylogeny.
MATERIALS AND METHODS
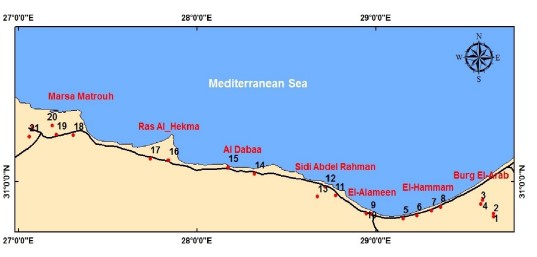
Study area: The studied area was the coastal strip of the Mediterranean Region, Alexandria, Egypt. The locations were demonstrated in Fig. 1. These locations are characterized by the salt marsh habitats and vegetation.
Materials: Field trips were carried out regularly, through three seasons; autumn, winter and spring; to the western coastal region of Egypt, from October, 2020 till April, 2022, to investigate the flowering stages of the halophytic species in this area. A survey of the salt habitats in the coastal region of Alexandria has been done to identify the common halophytic species in this region. Regions of collections illustrated in Fig. 1.
|
| Table 1: | Classifications of Chenopodiaceae s.l.6,7 and the studied species | |||
| Author | |||||
| Kuhn et al.7 | Hernández-Ledesma et al.6 | ||||
| Taxa Family |
Subfamily | Tribe | Subfamily | Tribe | Studied species |
| Amaranthaceae s.s | Amaranthoideae | Amarantheae | Amaranthoideae | Amarantheae | |
| Celosieae | Celosieae | ||||
| Gomphrenoideae | Bryolineae | Gomphrenoideae | Bryulineae | ||
| Gomphreneae | Gomphreneae | ||||
| Polycnemoideae | Polycnemeae | ||||
| Chenopodiaceae s.s | Chenopodioideae | Atripliceae | Betoideae | Beteae | |
| Beteae | Camphrosmoideae | Camphrosmeae | Bassia muricata | ||
| Camphorosmeae | Chenopodioideae | Atripliceae | Atriplex | ||
| Chenopodieae | Chenopodieae | Chenopodium album and C. murale | |||
| Corispermeae | Sclerolaeneae | ||||
| Sclerolaeneae | Corespermoideae | Corespermeae | |||
| Polycnemoideae | Polycnemeae | Salicornoideae | Halopeplideae | Arthrocnemum glaucum, | |
| Halocnemon strobilaceum | |||||
| and Salicornia fruticosa | |||||
| Salicornoideae | Halopeplideae | Salicomieae | Halopeplis amplexicaulis | ||
| Salicomieae | Salsoloideae | Salsoleae | Anabasis articulate, | ||
| Salsola inermis, | |||||
| S. kali, S. longifolia, | |||||
| S.tetragona and S. volkensii | |||||
| Salsoloideae | Salsoleae | Suaedoideae | Suaedeae | Suaeda fruticosa and | |
| Suaedeae | S. vermiculata | ||||
The recent taxonomic system gathered the six subfamilies and the fifteen tribes altogether under the family Chenopodiaceae s.l. Table 1 summarizes of the divisions of the Chenopodiaceae s.l.
The collected specimens were identified by the aid of the student’s flora of Egypt14,20 as well as by comparing the collected specimens with already identified herbarium specimens. Voucher specimens were kept in the Faculty of Science, Alexandria University, Botany and Microbiology Department Herbarium. Flowers of the collected taxa have been taken carefully for pollen grain examinations as follows:
Methods: Pollen grain samples of all studied taxa were acetolyzed according to Erdtman’s technique21. Slides were prepared from the acetolyzed portion of pollen grains for light microscope examination by mounting in glycerin jelly, examined and measured using a Zeiss light microscope with a pre-calibrated eyepiece micrometer. Measurements given are the means of 30 acetolyzed, well-developed pollen grains from each taxon.
| Table 2: | Studied taxa, collecting data, halophytic type, and their characteristic features | |||
| Studied taxa | Collecting data (Egypt) | Type of halophyte | Characteristic feature |
| Arthrocnemum glaucum (del.) Ung.-Sternb. | El-Omayed Road, El-Khashm Hill, | Obligate | Grow in saline habitats and increase in salt |
| N 30 48.293 E 029 11.485, 17.11.2016 | Concentration (≥ 200 mM) stimulate the growth | ||
| Halocnemum strobilaceum (Pallas) M. Bieb | El-Omayed Road, El-Khashm Hill, | ||
| Halopeplis amplexicaulis (Vahl) Ung.-Ster | N 30 48.293 E 029 11.485, 17.11.2016 | ||
| Salicornia fruticosa L. | |||
| Suaeda fruticosa (L.) Boiss | El-Hammam Road, Mariout Lake Sides, | ||
| Suaeda vermiculata Forssk. | kilo 65, N 30 52.866 E 029 22.170, 17.11.2016 | ||
| Atriplex halimus L. | Burg El-Arab Airport Road, km 52, | Facultative halophytes | Grow in moderate salt habitats (≤200 mM), but |
| N 30 56.519 E 092 30.145, 21.4.2016 | Optimum growth at lower salt concentrations | ||
| Anabasis articulata (Forssk.) Moq. | 95 km West of Alex; Alex-Matrouh Road Sides (Omayed), N 30 49.541 E 014 32.488, 16.10.2017 | ||
| Bassia muricata (L.) Murr. | Burg El-Arab Airport Road, | ||
| Chenopodium album L. | km 52, N 30 56.519 E 092 30.145, 21.4.2016 | ||
| Chenopodium murale L. | El-Hammam Road, Mariout Lake Sides, | ||
| Kilo 65, N 30 52.866 E 029 22.170, 17.11.2016 | |||
| Salsola inermis Forssk. | 80 km West of Alex; Alex-Matrouh Road Sides (Omayed), N 30 49.541 E 029 11.488, 16.10.2017 | Habitat-indifferent | Grow in salt free soil, but prefer salty habitats |
| Salsola kali L. Salsola longifolia Forssk. |
80 km west of Alex; Alex-Matrouh Road Sides (Omayed), N 30 49.541 E 029 11.488, 16.10.2017 | ||
| Salsola tetragona Del. | Burg El-Arab Airport Road, km 52, | ||
| N 30 56.519 E 092 30.145, 21.4.2016 | |||
| Salsola volkensii Asch. and Schweinf. | El-Omayed Road, El-Khashm Hill, | ||
| N 30 48.293 E 029 11.485, 17.11.2016 |
Non-acetolyzed pollen grains sputtered onto copper stubs, coated with 30 nm gold using fine coat ion sputter JEOL JFC 1100E, examined and photographed at 30 KV using JEOL JSM-3500 scanning electron microscope present in the Faculty of Science, Alexandria University. The terminology used in the present study is according to Larsen22 and Punt et al.23. The studied taxa, their taxonomical position, and their collecting data are summarized in Table 2.
RESULTS
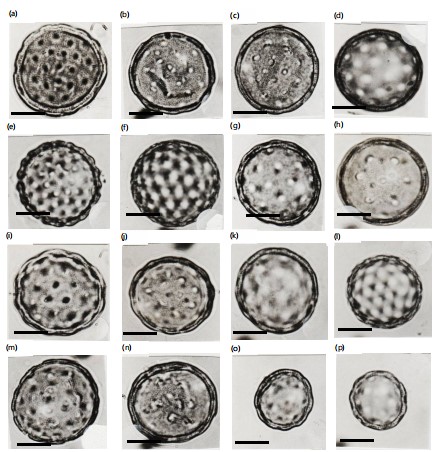
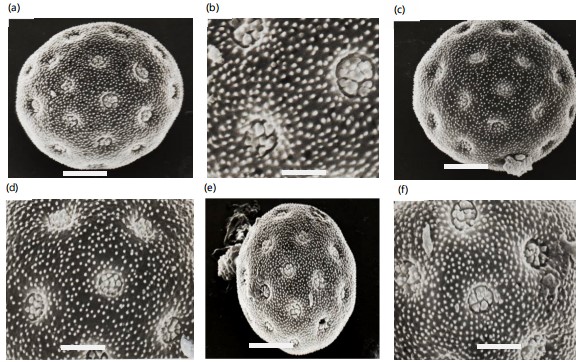
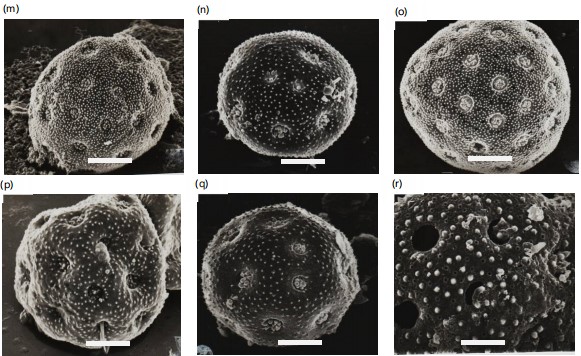
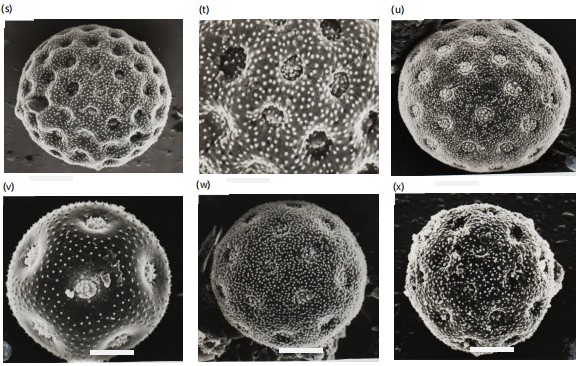
Light and scanning electron microphotographs were taken and listed in Fig. 2-3. Pollen morphological records are based on the grain size, aperture, number of pores, surface, and the exine of pollen grain (Fig. 2a-3b). The grains of the studied species of Chenopodiaceae are in both polar and equatorial views spheroidal with variable diameters. They are polypantoporate with a different number of pores, usually covered with dense granules or echinae. The exine is tectate with inperforate tectum and granulate or echinate surface. The thickness of the exine was usually thin, measured from 0.8 to 3.2 μm. The grain sizes are either small, 16.3 μm or large, 42.0 μm. According to variations in pollen diameters, three groups can be distinguished. Group 1 with a diameter 20 μm, group 2 with a diameter from 20-30 μm and group 3 with a diameter 30 μm. Pores are usually circular or eliptical with shiny edges or a granulated band. The size of pores is 0.5 to 3.2 μm. The number of pores are between 12 and reaches 62 (Table 3). According to the pollen diameter, three main groups can be recognized: Small pollen grains with a diameter never exceed 20 μm and have three Salsola species only, S. longifolia, S. tetragona, and S. volkensii (group 1). The moderate pollen grains with a diameter of 20-30 μm have six taxa; Atriplex halimus, Anabasis articulata,
Chenopodium album, C. murale, Salsdola innermis, and S. kali (group 2) and the third group (group 3) has seven taxa, all the obligate halophytes, in addition to Basia muricata. These three groups can be divided into three subgroups according to the number of pores. Subgroup 1 with pollen grains have fewer than 20 pores/pollen, subgroup 2 has pollen grains with more than 20 and fewer than 40 pores/pollen, while subgroup 3 has pollen grains with more than 50 pores/pollen. Pollen of each species has been described in characteristics of a size, aperture, pore size and number, and exine thickness and ornamentation, and documented with SEM photographs (1-16). Detailed descriptions of the pollen characters of each species, arranged according to their taxonomic positions listed in Table 2. The different types of halophyte mentioned in this work are according to their ecology as mentioned in an earlier study by Grigore and Toma13.
Obligate halophytes
Arthrocnemum glaucum: Pollen grains radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter 40.2 μm in equatorial view, pores moderate, deeply sunken from 3.2-4.1 μm in diameter. the surrounding rim is narrow and faintly granular, interporal distance of 3.2-4.8 μm, and the ratio between the interporal distance to the pore diameter from 1.2-1.8. The amount of pores/pollen from 30-38. Exine thin 2.0-2.4 μm tectate with echinate surface (Fig. 2a, 3a-b).
Halocnemum strobilaceum: Pollen grains radially symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 36.4 μm in equatorial view, pores moderate, superficial from 2.4-3.6 μm in diameter, the surrounding rim is narrow and smooth, interporal distance from 2.8-3.6 μm, and the ratio between the interporal distance to the pore diameter from 1.0-1.7. The amount of pores/pollen from 26-32. Exine moderate in thickness, 2.2-2.8 μm tectate with granulate surface (Fig. 2b, 3c-d).
Halopeplis amplexicaulis: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 35.4 μm in equatorial view, pores moderate, superficial from 2.0-2.8 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 3.4-4.6 μm, and the ratio between the interporal distance to the pore diameter from 1.6-1.7. The number of pores/pollen from 22-28. Exine thin 2.2-2.8 μm tectate with granulate surface (Fig. 2c, 3e-f).
| Table 3: | Pollen morphological characters in the studied species |
|||
| Character | |||||||||||||
| Pore | Exine | ||||||||||||
| Species | Di (Mean±SD) |
State | D (μm) |
IP (C) (μm) |
C/D (μm) |
No. | DG./P (μm) |
Th (μm) |
Or (μm) |
DG (μm) |
G | SG | HT |
| Arthrocnemum glaucum | 40.2±0.4 | S. | 3.2-4.1 | 4.0±0.36 | 1.2-1.8 | 35±0.7 | +++ | 2.0-2.4 | Ec | ++ | G3 | SG 2 | Obligate |
| Halocnemon strobilaceum | 36.4±0.2 | Sup. | 2.4-3.6 | 3.2±0.22 | 1.0-1.7 | 32±0.2 | ++ | 2.2-2.8 | Gr | ++ | |||
| Halopeplis amplexicaulis | 35.4±0.3 | Sup. | 2.0-2.8 | 4.0±0.4 | 1.6-1.7 | 26±0.5 | +++ | 2.2-2.8 | Ec | ++ | |||
| Salicornia fruticosa | 35.2±0.6 | S. | 0.8-1.2 | 2.8±0.2 | 2.7-3.1 | 56±0.2 | ++ | 1.8-2.4 | Gr | +++ | SG 3 | ||
| Suaeda fruticosa | 32.2±0.5 | Sup. | 1.0-1.2 | 2.2±0.4 | 1.8-2.2 | 58±0.4 | ++ | 2.4-3.0 | Gr | +++ | |||
| Suaeda vermiculata | 35.8±0.6 | Sup. | 1.2-1.6 | 2.4±0.3 | 1.5-2.0 | 56±0.2 | ++ | 2.4-3.2 | Gr | +++ | |||
| Atriplex halimus | 27.9±0.2 | D.S. | 0.8-1.0 | 4.70±0.3 | 5.8-6.3 | 36±0.4 | +++ | 1.0-1.3 | Gr | +++ | G2 | SG2 | Facultative |
| Anabasis articulata | 25.8±0.8 | S. | 2.8-4.0 | 4.9±0.3 | 1.3-1.6 | 16±0.6 | +++ | 2.6-3.2 | Gr | +++ | SG1 | halophytes | |
| Bassia muricata | 42.0±0.8 | S. | 0.5-0.9 | 4.2±0.2 | 5.4-5.5 | 52±0.8 | + | 1.9-2.1 | Gr | + | G3 | SG 3 | |
| Chenopodium album | 26.9±0.4 | Sup. | 0.8-0.9 | 4.2±0.34 | 4.4-4.9 | 24±0.7 | +++ | 1.2-1.5 | Ec | +++ | G2 | SG 2 | |
| Chenopodium murale | 27.2±0.2 | S. | 0.8-0.9 | 4.4±0.14 | 4.5-4.8 | 26±0.4 | +++ | 1.6-1.9 | Ec | +++ | |||
| Salsola inermis | 20.7±0.2 | D.S. | 1.6±0.2 | 1.3±0.2 | 0.7-0.8 | 67±0.8 | ++ | 2.2-2.4 | Gr | ++ | SG 3 | Habitat- | |
| Salsola kali | 24.8±0.2 | D.S. | 1.3±0.1 | 1.6±0.1 | 1.2-1.3 | 52±0.5 | +++ | 2.3-2.4 | Ec | ++ | indifferent | ||
| Salsola longifolia | 19.8±0.2 | S. | 1.3±0.1 | 2.0±0.2 | 1.5-1.6 | 12±0.2 | +++ | 2.4-2.7 | Gr | ++ | G 1 | SG 1 | |
| Salsola tetragona | 18.8±0.2 | D.S. | 0.7±0.1 | 1.3±0.1 | 2.1-2.2 | 28±0.4 | +++ | 2.0-2.6 | Gr | ++ | SG 2 | ||
| Salsola volkensii | 16.3±0.2 | D.S. | 1.1±0.1 | 1.1±0.1 | 1-1.2 | 32±0.2 | +++ | 2.4-2.6 | Gr | +++ | |||
| C/D: Interporal distance/pore diameter, D: Pore diameter, DG: Density of granules or echinae, Di: Diameter, D.G./P: Density of granules over pore, +: Sparse, ++: Moderate, +++: Dense (crowed), D.S: Deeply sunken, Ec: Echinate, G: Group, G1: Diameter 20 μm, G2: Diameter from 20-30 μm, G3: Diameter 30 μm Gr: Granulate, HT: Halophytic type, IP (C): Interporal distance, No: Number, Or: Ornamentation, S: Sunken, SG: Subgroup, Sup: Superficial and Th: Thickness | |||||||||||||
|
Salicornia fruticosa: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 35.2 μm in equatorial view, pores small, slightly sunken from 0.8-1.2 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 2.5-3.2 μm, and the ratio between the interporal distance to the pore diameter from 2.7-3.1. The amount of pores/pollen from 48-60. Exine thin 1.8-2.2 μm tectate with dense granulate surface (Fig. 2d, 3g-h).
Suaeda fruticosa: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 32.2 μm in equatorial view, pores small, superficial from 1.0-1.2 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 2.0-2.2 μm, and a ratio between the interporal distance to the pore diameter from 1.8-2.2. The number of pores/pollen from 48-60. Exine thin 2.4-3.0 μm tectate with densely granulate surface (Fig. 2e, 3i-j).
Suaeda vermiculata: Pollen grains radially symmetrical, apolar, pantoporate, spherical, mean pollen diameter 35.8 μm in equatorial view, pores small, superficial from 1.2-1.6 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 2.2-2.7 μm, and the ratio between the interporal distance to the pore diameter from 1.5-2.0. The number of pores/pollen from 48-58. Exine thin 2.4-3.2 μm tectate with densely granulate surface (Fig. 2f, 3k-l).
Facultative halophytes
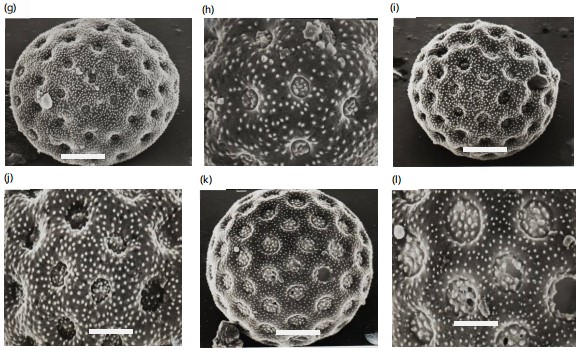
Atriplex halimus: Pollen grains radially symmetric, apolar, pantoporate, spheroidal or rarely subprolate, mean pollen diameter 27.9 μm in equatorial view. Pores are small, deeply sunken from 0.8-1.0 μm, without any surrounding rim, and interporal distance from 4.4-5.2 μm. The ratio between the interpore distance to the pore diameter from 5.8-6. The average number of pores/pollen is 36, which are covered by dense granules. The exine is thin from 1.0-1.3 μm, tectate with dense granulate surface (Fig. 2g, 3m).
|
Anabasis articulate: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 25.8 μm in equatorial view. Pores are moderate, sunken in the middle of pentagonal exinous elevations, from 2.8-4.0 μm in diameter. The surrounding rim is undistinguishable, interporal distance from 4.6-5.2 μm, and the ratio between the interporal distance to the pore diameter from 1.3-1.6. The amount of pores/pollen from 12-20. Exine moderate 2.6-3.2 μm tectate with densely granulate surface (Fig. 2h, 3n).
Bassia muricata: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 42.32 μm in equatorial view. pores are small, slightly sunken from 0.5-0.9 μm in diameter. The surrounding rim is narrow and smooth, interporal distance of 3.8-4.4 μm, and the ratio between the interporal distance to the pore diameter from 5.4-5.5. The number of pores/pollen from 45-58. Exine thin 1.9-2.1 μm tectate with sparse granulate surface (Fig. 2i, 3o).
Chenopodium album: Pollen grains radially symmetric, apolar, pantoporate, spheroidal, mean pollen diameter 26.9 μm in equatorial view. Pores are small, superficial, ranging from 0.8-0.9 μm, the surrounding rim is narrow and faint and covered by dense granules, interporal distance from 3.9-4.8 μm, and the ratio between the interporal distance to the pore diameter from 4.4-4.9. The number of pores/pollen from 18-28. Exine thin 1.2-1.5 μm tectate with dense echinate surface (Fig. 2j, 3p).
Chenopodium murale: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter 27.2 μm in equatorial view, pores small, slightly sunken from 0.8-0.9 μm in diameter, the surrounding rim is narrow and smooth, interporal distance of 3.9-4.8 μm and the ratio between the interporal distance to the pore diameter from 4.2-5.0. The number of pores/pollen from 23-28. Exine thin 1.6-1.9 μm tectate with dense echinate surface (Fig. 2k, 3q-r).
Habitat indifferent
Salsola inermis: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 20.7 μm in equatorial view, pores small, deeply sunken from 1.4-1.8 μm in diameter, the surrounding rim is narrow, interporal distance from 1.0-1.5 μm, and the ratio between the interporal distance to the pore diameter from 0.7-0.8 The number of pores/pollen from 50-67. Exine thin 2.2-2.4 μm tectate with moderately granulate surface (Fig. 2l, 3s-t).
Salsola kali: Pollen grains are radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter of 24.8 μm in equatorial view. pores moderate, deeply sunken from 1.2-1.5 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 1.5-1.8 μm, and the ratio between the interporal distance to the pore diameter from 1.2-1.3. The number of pores/pollen from 40-53. Exine thin 2.3-2.4 μm tectate with moderately echinate surface (Fig. 2m, 3u).
Salsola longifolia: Pollen grains are radially symmetrical, apolar, pantoporate spherical, mean pollen diameter of 19.8 μm in equatorial view, pores moderate, sunken from 1.2-1.5 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 1.8-2.2 μm, and the ratio between the interporal distance to the pore diameter from 1.5-1.6. The number of pores/pollen from 12-15. Exine thin 2.4-2.7 μm tectate with moderately granulate surface (Fig. 2n, 3v).
Salsola tetragona: Pollen grains radial symmetrical, apolar, pantoporate, spherical, mean pollen diameter 18.8 μm in equatorial view, pores small, deeply sunken from 0.6-0.8 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 1.8-2.2 μm and the ratio between the interporal distance to the pore diameter from 2.1-2.2 μm. The number of pores/pollen from 25-30. Exine thin 2.0-2.6 μm tectate with moderately granulate surface (Fig. 2o, 3w).
Salsola volkensii: Pollen grains radially symmetrical, apolar, pantoporate, spherical, mean pollen diameter 16.3 μm in equatorial view, pores small, deeply sunken from 0.8-1.3 μm in diameter, the surrounding rim is undistinguishable, interporal distance from 1.0-1.5 μm, and the ratio between the interporal distance to the pore diameter from 1.0-1.2. The number of pores/pollen from 28-35. Exine thin 2.4-2.6 μm tectate with densely granulate surface (Fig. 2p, 3x).
DISCUSSION
Pollen morphology has been considered an important tool in taxonomic treatments of several taxa for a long time ago. Palynological studies have been used in taxonomic treatments of the family Chenopodiaceae9-12,24. Perveen and Qasier25 studied the pollen grains morphology of the Pakistani flora by both light and scanning electron microscopes. They described the Chenopodiaceae family by being a stenopalynous family. They found that pollens are usually radially symmetrical, apolar, pantoporate, and spheroidal. This finding coordinates with the results obtained by Tsymbalyuk et al.26 and the data obtained in this investigation. The three categories of halophytes have apolar spheroidal pollen grains with zonoporate apertures. The apertures are covered by granulated or echinate exinous surface in different densities. Lu et al.27 recorded six pollen types within the taxa of Chenopodiaceae grown in the Eastern Central Asia. They linked the six types to the different plant communities which dominated by chenopods taxa. These pollen types were according to number of pores, pore diameters and density of granules over the pores and referred the six groups to the different plant communities in the xerophytic deserts. Our results are in partial improvement with Lu et al.27 conclusion and coincide with Jalilzadeh et al.28 results, who recorded three pollen types in the Iranian Chenopodiaceae coordinate with the recent taxonomic treatment of the family; group 1 has the Atriplex taxa, group 2 has the Chenopodium taxa, while group 3 has the Dysphania taxa. These groups were according to pore characters as well. In the studied taxa the number of pores varied greatly from less than 20 pore/pollen in both Anabasis articulata and Salsola longifolia to more than 60 pore/pollen in Salsola innermis. In the rest of the studied taxa the pores varied greatly in numbers. These variations in the number of pores did not associate with the three investigated halophyte types, but it may be in relation to the degree of soil salinity. Meanwhile, all the studied taxa have pollen grains with pores covered with either dense or moderate granules or echinae. The density of the granules over the pores was independent of the halophytic type.
In general, the studied pollen taxa can be classified under three categories according to their number of pores/pollen, the first one with taxa having pollen grains with pores less than 20, which includes both Anabasis articulata and Salsola longifolia. The second category with taxa having pollen grains with pores from 20-40 and includes eight taxa: Arthrocnemum glaucum, Halocnemum strobilaceum, Halopeplis amplexicaulis, Atriplex halimus, Chenopodium album, C. murale, Salsola tetragona and S. volkensii. The third category gathered the rest of the studied taxa with several pores/pollen more than 50 pores. These groupings coordinate with those obtained by Jalilzadeh et al.28. But when considering the size of the pollen grain diameter, another three groupings can be realized. The small pollen grains with a diameter never exceeding 20 μm have three Salsola species only, S. longifolia, S. tetragona, and S. volkensii. The moderate pollen grains with a diameter from 20-30 μm have six taxa: Atriplex halimus, Anabasis articulata, Chenopodium album, C. murale, Salsola innermis, and S. kali. The third group has seven taxa, all the obligate halophytes in addition to Basia muricata. These three groups can be divided into three subgroups according to the number of pores. Subgroup 1 with pollen grains have fewer than 20 pores/pollen, subgroup 2 has pollen grains with more than 20 and fewer than 40 pores/pollen, while subgroup 3 has pollen grains with more than 50 pores/pollen. This categorization can be considered most reliable and coincides with that of Sonyan et al.29, and classified the Trans-caucasian taxa of subfamily Salsoloideae in two pollen morphological types and four subtypes. The subdivision of the subtypes was according to differences in the size of pollen grains. The data obtained in this work in agreement with those obtained by Nazish et al.30,31 as the size of the pollen grains is slightly different between the taxa, but their general pollen features are common between the halophytic species. This study recommends the pollen morphological features such as shape, size, exine sculpture, and aperture number and type useful reference for the systematic classification of halophytes, as recommended by Nazish et al.31 .
CONCLUSION
Pollen morphology is considered one of the important taxonomic tools in plant identification and taxa discrimination. The current work confirms the importance of the light and scanning electron microscope examinations in understanding the major pollen features of halophytes of Chenopodiaceae taxa. The results obtained illustrate the common pollen features of the three halophytic groups. These common features can be an indication of their common ancestors and are subject to different evolutionary lines. This work clarifies the similarities and differences in the pollen characters of the studied halophytic taxa from the family Chenopodiaceae from the salt habitats in the western coastal region of Egypt.
SIGNIFICANCE STATEMENT
Halophytic plants exhibit various adaptive mechanisms to survive in high-salinity environments, one of which involves modifications in pollen morphology and internal tissues. This study aims to investigate the pollen morphological characteristics of halophytic species within the family Chenopodiaceae to understand their potential adaptive significance and evolutionary relationships. According to pollen diameters followed by the number of pores/pollen, three groups have been identified, with three subgroups. This study clarified that the halophytes may be of the same ancestral origin, which are subject to different evolutionary lines.
REFERENCES
- Garcia-Caparros, P., M.J. Al-Azzawi and T.J. Flowers, 2023. Economic uses of salt-tolerant plants. Plants, 12.
- Mann, A., C. Lata, N. Kumar, A. Kumar, A. Kumar and P. Sheoran, 2023. Halophytes as new model plant species for salt tolerance strategies. Front. Plant Sci., 14.
- Öztürk, M., V. Altay, M. Nazish, M. Ahmad and M. Zafar, 2023. Halophyte Plant Diversity and Public Health. 1st Edn., Springer International Publishing, Switzerland, ISBN: 978-3-031-21944-3, Pages: 203.
- Wanas, A.L., A.S. Abd El-Hamied, E.M. Shabka and M.S. Serag, 2018. A survey study on some halophytes growing in the Damietta coastal area referring to its botanical and ecological significance. J. Plant Prod., 9: 641-648.
- Bazos, I., I.P. Kokkoris and P. Dimopoulos, 2021. Diversity of Halophytes and Salt Tolerant Plants at the Species-, Habitats- and High-Rank Syntaxa Level in Greece. In: Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture, Grigore, M.N. (Ed.), Springer, Cham, Switzerland, ISBN: 978-3-030-57635-6, pp: 787-820.
- Hernández-Ledesma, P., W.G. Berendsohn, T. Borsch, S. von Mering and H. Akhani et al., 2015. A taxonomic backbone for the global synthesis of species diversity in the angiosperm order Caryophyllales. Willdenowia, 45: 281-383.
- Kühn, U., V. Bittrich, R. Carolin, H. Freitag, I. C. Hedge, P. Uotila and P.G. Wilson, 1993. Chenopodiaceae. In: Flowering Plants · Dicotyledons: Magnoliid, Hamamelid and Caryophyllid Families, Kubitzki, K., J.G. Rohwer and V. Bittrich (Eds.), Springer, Berlin Heidelberg, ISBN: 978-3-662-02899-5, pp: 253-281.
- Fuentes-Bazan, S., P. Uotila and T. Borsch, 2012. A novel phylogeny-based generic classification for Chenopodium sensu lato, and a tribal rearrangement of Chenopodioideae (Chenopodiaceae). Willdenowia, 42: 5-24.
- Flowers, T.J. and T.D. Colmer, 2008. Salinity tolerance in halophytes. New Phytol., 179: 945-963.
- Barakat, N.A.M., E. Cazzato, B. Nedjimi, H.F. Kabiel, V. Laudadio and V. Tufarelli, 2014. Ecophysiological and species-specific responses to seasonal variations in halophytic species of the chenopodiaceae in a Mediterranean salt marsh. Afr. J. Ecol., 52: 163-172.
- Waldron, B.L., J.K. Sagers, M.D. Peel, C.W. Rigby, B. Bugbee and J.E. Creech, 2020. Salinity reduces the forage quality of Forage kochia: A halophytic Chenopodiaceae shrub. Rangeland Ecol. Manage., 73: 384-393.
- Song, X., Y. Su, J. Zheng, Z. Zhang, Z. Liang and Z. Tang, 2022. Study on the effects of salt tolerance type, soil salinity and soil characteristics on the element composition of chenopodiaceae halophytes. Plants, 11.
- Grigore, M.N. and C. Toma, 2017. Definition and Classification of Halophytes. In: Anatomical Adaptations of Halophytes: A Review of Classic Literature and Recent Findings, Grigore, M.N. and C. Toma (Eds.), Springer, Cham, Switzerland, ISBN: 978-3-319-66480-4, pp: 3-28.
- Tackholm, V., 1974. Students Flora of Egypt. 2nd Edn., Cairo University Cooperative Printing Co., Cairo, Egypt, Pages: 888.
- Olvera, H.F., S. Fuentes-Soriano and E.M. Hernandez, 2006. Pollen morphology and systematics of Atriplex (Chenopodiaceae). Grana, 45: 175-194.
- Dehghani, M. and H. Akhani, 2009. Pollen morphological studies in subfamily Suaedoideae (Chenopodiaceae). Grana, 48: 79-101.
- Bahadur, S., M. Ahmad, S. Mir, M. Zafar, S. Sultana, S. Ashfaq and M. Arfan, 2018. Identification of monocot flora using pollen features through scanning electron microscopy. Microsc. Res. Tech., 81: 599-613.
- Khan, R., S. Zain Ul Abidin, M. Ahmad, M. Zafar, J. Liu and H. Amina, 2018. Palyno-morphological characteristics of gymnosperm flora of Pakistan and its taxonomic implications with LM and SEM methods. Microsc. Res. Tech., 81: 74-87.
- Dehghani, M. M. Djamali and H. Akhani, 2021. Pollen morphology of the subfamily Salicornioideae (Chenopodiaceae) in Eurasia and North Africa. Palynology, 45: 245-258.
- Shaltout, K.H., 2004. An updated flora of Egypt. Divers. Distrib., 10: 77-78.
- Tsymbalyuk, Z.M., S. Çelenk, C.D. Bell, L.M. Nitsenko and S.L. Mosyakin, 2022. Comparative palynomorphological study of the genus Symphoricarpos (Caprifoliaceae): Exine sculpture and implications for evolution. Palynology, 46: 1-14.
- Larsen, S.S., 1974. Pollen morphology of Thai species of Bauhinia (Caesalpiniaceae). Grana, 14: 114-131.
- >Punt, W., P.P. Hoen, S. Blackmore, S. Nilsson and A. le Thomas, 2007. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol., 143: 1-81.
- Turki, Z., F. El-Shayeb and F. Shehata, 2008. Taxonomic studies in the Camphorosmeae (Chenopodiaceae) 1. Subtribe: Kochiinae (Genera: Bassia All., Kochia Roth and Chenolea Thunb.). Acta Bot. Hung., 50: 181-201.
- Perveen, A. and M. Qasier, 2012. Pollen flora of Pakistan-LXX: Chenopodiaceae. Pak. J. Bot., 44: 1325-1333.
- Tsymbalyuk, Z.M., D. Ivanova and L.M. Nitsenko, 2024. Comparative morphometric and morphological study of the pollen of Beta trigyna, B. vulgaris and B. vulgaris subsp. maritima (Chenopodiaceae/Amaranthaceae sensu APG IV). Hacquetia, 23: 239-252.
- Lu, K.Q., M. Li, G.H. Wang, L.S. Xu and D.K. Ferguson et al., 2019. New pollen classification of Chenopodiaceae for exploring and tracing desert vegetation evolution in Eastern Arid Central Asia. J. Syst. E, 57: 190-199.
- Jalilzadeh, A., S.M.M. Hamdi, Y. Asri, M. Assadi and A. Iranbakhsh, 2021. Pollen micromorphological study of some Chenopodiaceae species and their taxonomical relationships in Iran. Rostaniha, 22: 209-222.
- Sonyan, H., A. Hayrapetyan, N. Poghosyan and A. Muradyan, 2022. Pollen typification and palynotaxonomy of subfamily Salsoloideae Ulbr. (Chenopodiaceae Vent.) in the flora of South Transcaucasia. Acta Palaeobot., 62: 93-107.
- Nazish, M., F. Bibi, M.S. Alwahibi, M.S. Elshikh and W. Zaman, 2024. Exploration of salinity effect on palyno-morphological characteristics of plant species collected from salt range. Bangladesh J. Plant Taxon., 31: 33-49.
- Nazish, M., A. Kamal, M.A. Khan, Khursheed Ur Rahman and M.S. Elshikh et al., 2025. Integration of multivariate indices to correlate pollen morphology and evolutionary adaptations in halophytes. Genet. Resour. Crop E.
How to Cite this paper?
APA-7 Style
Taia,
W.K. (2025). Comparative Analysis of Pollen Morphological Traits in Halophytic Chenopods. Research Journal of Botany, 20(1), 191-202. https://doi.org/10.3923/rjb.2025.191.202
ACS Style
Taia,
W.K. Comparative Analysis of Pollen Morphological Traits in Halophytic Chenopods. Res. J. Bot 2025, 20, 191-202. https://doi.org/10.3923/rjb.2025.191.202
AMA Style
Taia
WK. Comparative Analysis of Pollen Morphological Traits in Halophytic Chenopods. Research Journal of Botany. 2025; 20(1): 191-202. https://doi.org/10.3923/rjb.2025.191.202
Chicago/Turabian Style
Taia, Wafaa, Kamal.
2025. "Comparative Analysis of Pollen Morphological Traits in Halophytic Chenopods" Research Journal of Botany 20, no. 1: 191-202. https://doi.org/10.3923/rjb.2025.191.202

This work is licensed under a Creative Commons Attribution 4.0 International License.