Leaf Morphological Variations Among Selected Bauhinia L. Species
-
Wafaa Kamal Taia

Department of Botany and Microbiology, Faculty of Science, Alexandria University, Alexandria, Egypt
Eman Mohammed BassioniDepartment of Botany and Microbiology, Faculty of Science, Alexandria University, Alexandria, Egypt
| Received 01 May, 2025 |
Accepted 29 Jun, 2025 |
Published 30 Jun, 2025 |
Background and Objective: Bauhiniatrees and lianas have been used in many traditional medicines for their richness in natural products. Many of its species need accurate naming and description to avoid misidentification. This study investigates leaf variations in Bauhinia species. Materials and Methods: Leaf macro- and micro-morphological characters of eleven Bauhinia species commonly planted in the Egyptian streets were examined and measured using an eyepiece and both light and scanning electron microscopes. Different leaf shapes, epidermal cells, hair, and stomatal characters were recorded. Results: Variations in epidermal cell shapes, periclinal and anticlinal walls, besides hair density, type, and their wall ornamentation were highly different. Type of stomata, stomatal index, and density were valuable characters in the differentiation of the studied species. According to SEM examination, most of the studied species have epicuticular wax depositions with different shapes. The most important characters in discriminating the studied taxa are their macro-morphological ones, besides hair and stomatal characters. Conclusion: This study supports the taxonomical division of the Bauhinia species into two subgenera with five sections. This study can aid in tracing the adaptation strategy of the different species to the polluted environment in Egypt. Identification key, adaptation strategy, and postulated evolutionary groupings within the studied species according to leaf macro-morphological characters were given.1-B. variegatais considered primitive, while B. Grevei was considered the most advanced species.
| Copyright © 2025 Taia and Bassioni. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Genus Bauhinia is considered one of the widely used genera in traditional medicine. Its species are rich in natural compounds that can be used to cure various diseases1 . Species identification and description are considered an important aim to avoid misidentification of the medicinally used species. Bauhinia L. trees are ornamental trees planted in most of the Egyptian streets. The legume phylogeny working group2considered all the Bauhinia trees as a single genus, which was subdivided into several subgenera. The genus is considered one of the largest and diverse genera under the family Fabaceae, subfamily Cercidoideae, tribe Bauhinieae, subtribe Bauhiniinae. It comprises about 300 species with a wide distribution in the tropical and subtropical regions. The genus name, Bauhinia, was given after the two brothers Jean and Gaspard Bauhin, according to the bilobate leaves of the genus3. The classification of the Bauhinia species has been subjected to many scientific opinions3-8. All the species have common bilobate leaves with different sizes and apical notch depths, and shapes. These species were grouped under four subgenera by Duarte-Almeida et al.9, Barklya (1 species), Bauhinia (140 species), Elayuna (6 species), and Phanera (150 species); according to their life forms, while6,10 treated all the Bauhinia trees as one large genus, considering the variations in its morphological characters as subgenera. Igbari et al.11, Trethowan et al.12 and Meng et al.13 separated the Bauhinia species into eight to nine genera based on their morphological variations. All these taxonomic classifications of Bauhinia were dependent on the leaf morphological variations, fruit characters, and habit of the plant. Tucker14 considered the Bauhinia species as a natural, evolutionary unit with a reticulate pattern of variations across the pantropic range.
Leaf characters are considered from the first steps in taxonomical keys and help in understanding the adaptation strategy of the plants15. Hickey16 mentioned that the neglect of leaf macro-morphological characters in the taxonomical comparison of the taxa was due to the lack of detailed classification of their features. Afterwards, taxonomists realized the importance of the accurate study of the leaf features as an identification tool and to understand the adaptation strategy of the plants17-19. Modern plant classification based on molecular techniques and molecular analyses has the leaves an important role in scientific research and new classifications. These new classifications, based on leaf extracts and molecular analyses and neglecting the morphological characters of the leaves, resulted in many unreliable statuses within many taxa. Accordingly, the macro and micro-morphological leaf characters, besides many other morphological characters, became overlooked in taxonomical works. Smykal et al.20 used the variability in foliar epidermal micromorphology within 5 species of Bauhinia to determine the suitability for interspecies taxonomic delimitation and identification. Feodorova and Alexandrov21 found that the use of leaf morphological characters was diagnostic in the differentiation between black and white poplars. While El-Gazzar and Moustafa22 found that leaf characters were useful tools in discriminating the Egyptian Solanum species. According to the use of leaf macro-morphological characters in the discrimination of the families, Taia and El-Badan23 found that the leaf characters confirmed the separation of the two families as separate families. Taia et al.24 used the leaf characters in the identification and separation of the species under the tribe Malave. The useful work in the delimitation of the monocot orders using leaf characters was that done by Taia et al.25 as they found that characters of epidermal cells, hairs, papillae, prickles, and stomata can give clear features in the identification of some species.
Meanwhile, the leaf characters can indicate the adaptation strategy of the plants. Yao-Qi and Zhi-Heng15, Taia et al.19 indicated the importance of studying leaf morphological characters as leaf size, type, shape, margin type, besides several teeth and all the leaf morphological features, to understand their adaptive strategies in obtaining their resources. They indicated the importance of hair density, leaf texture, and stomatal density as ways of adaptation in the polluted areas.
The different species of genus Bauhinia were chosen for this study for their characteristic leaves and to clarify the variations in both the macro and micro-characters between its species. This study has been carried out to assess its taxonomic division and species identification.
MATERIALS AND METHODS
Study area: This study deals with eleven species commonly planted in the Egyptian streets and gardens as ornamental trees. The study focused on herbarium sheets and fresh materials gathered from Alexandria city streets. The specimens have been collected from August to November, 2022 and 2023. Examination of the herbarium sheets from the listed botanical gardens in Cairo, Egypt (Table 1) was carried out and measured. Well-representative leaves from at least 15 branches were examined carefully by eye lens and stereomicroscope to observe the leaf surfaces and texture. At least ten leaves were measured by ruler for their lengths and width (widest part) and L/W calculated. Leaf morphological characters based on both eye-lens and stereomicroscope examinations on ten mature leaves, photographed using an ordinary camera. Terminology used in the description of the morphological characters is that of Stearn26.
| Table 1: | Studied species of Bauhinia, source of materials, and confirmation of nomenclature | |||
| Bauhinia acuminata L. | Giza: Mazhar botanical garden, Egypt | ILDIS, USDA-ARS, IPNI |
| Bauhinia×blakeana Dunn | The national flowers of Hong Kong | |
| Bauhinia forficata Link | ||
| Bauhinia galpinii N.E. Br. | Al-Abeed agriculture farm, Egypt | |
| Bauhinia grevei Drake | Giza: Mazhar botanical garden, Egypt | |
| Bauhinia madagascariensis Desv. | ILDIS,USDA-ARS, IPNI | |
| Bauhinia monandra Kurz | Giza: Orman botanical garden, Egypt | |
| Bauhinia purpurea L. | ||
| Bauhinia tomentosa L. | Giza: Mazhar botanical garden, Egypt | |
| Bauhinia vahlii Wight & Arn L. | ||
| Bauhinia variegata L. | Plants of the world online |
Stomata: Leaf peels from both the abaxial and adaxial surfaces, at least five leaves, stained with safranin, mounted in a few drops of water and glycerol on clean microscopic slides and examined carefully by a 10×40x compound light microscope to observe the stomatal type, shape of epidermal cells, and hair density.
The stomata index (I) and density (D) were calculated by the expressions:
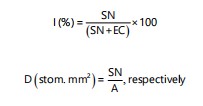 |
Where:
| SN | = | Stomata number | |
| EC | = | Epidermal cells number | |
| A | = | Area (mm2) |
Terminology used in the description of the morphological characters is that of Dilcher27.
SEM investigation: After the first examinations, parts of the dry leaves were put onto labeled Aluminum stubs furnished by double cello tape. The stubs were coated with 30 nm gold in a Polaron JFC-1100E coating unit, and examined carefully, photographed under 15 kV, with a JEOL JSM-IT200 SEM in the Electron Microscopes Unit, Faculty of Science, Alexandria University, Egypt. Terminology used according to Punt et al.28.
RESULTS
Macro-morphology: Most of the studied taxa are trees or shrubs; lianas are found only in Bauhinia vahlii. All species possess stipulate leaves. In B. blakeana, B. galpinii, B. grevei, B. madagascariensis, and B. purpurea, the stipules are adnate to the stem, forming protective pockets around the axillary buds. In contrast, B. acuminata, B. monandra, B. tomentosa, B. vahlii, and B. variegata exhibit free stipules, which appear as small, filamentous, leafy appendages. In B. forficata, the stipules are small, hard, and spine-like, with two sharp projections at the base of the leaf.
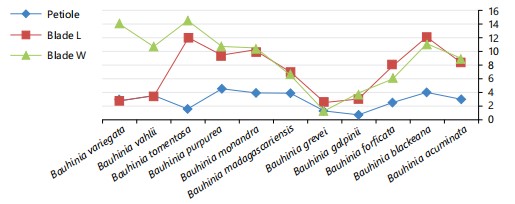
All examined taxa have petiolate leaves with varying petiole lengths (Fig. 1). Considerable variability is observed in leaf blade morphology and measurements among the studied species (Fig. 1). The leaf blades are generally simple with orbicular or cordate shapes, except in B. glabra and B. grevei, where the leaves are bifoliate and either oblong or nearly ovate. The leaf texture ranges from papyraceous, coriaceous, to scarious, whereas in B. vahlii, it is spongy with a soft, cushion-like feel (Table 2, Fig. 3a-k).
|
|
| Table 2: | Leaf macro-morphological characters of the studied species | |||
| Leaf blade | |||||||||
| Taxa | LF | St | F | B | S | T | Sf | V | Ap |
| Bauhinia acuminata | Sh | Fr | Si | St | Ob | Cor | Re | PE | Vs |
| Bauhinia blakeana | Tr | Ad | Si | Cv | Co | Sc | Cu | PE | Wv |
| Bauhinia forficata | Tr | Sp | Si | St | Co | Pa | Nr | PE | Y |
| Bauhinia galpinii | Sh | Ad | Si | Cv | Co | Sc | Nr | PE | Wv |
| Bauhinia greveri | Sh | Ad | Bl | St | Ob | Pa | Nr | UB | Ac |
| Bauhinia madagascariensis | Tr | Ad | Si | St | Or | Pa | Ner | PE | NV |
| Bauhinia monandra | Sh | Fr | Si | St | Or | Pa | Nr | PB | Vs |
| Bauhinia purpurea | Tr | Ad | Si | Cv | Or | Cor | Cu | PE | Vs |
| Bauhinia tomentosa | Tr | Fr | Si | St | Ob | Sp | Re | PE | Vs |
| Bauhinia vahlii | L | Fr | Si | Cv | Ob | Cor | Nr | PE | Y |
| Bauhinia variegata | Tr | Fr | Si | Cv | Ob | Cor | Cu | PE | NV |
| Al: Allover, An: Anisocytic, Ao: Anomocytic, AW: Anticlinal wall, Cv: Convex, D: Dense, Fl: Flate, D: Density, Ep: Epidermal, Gl: Globular, Gr: Granulate, M: Margin, Mo: Moderate, MUP: Multicellular uniseriate glandular, MUT: Multicellular uniserriate tabular, P: Position, Pa: Paracytic, PW: Periclinal wall, S: Superficial, Sc: Scally, Si: Sinuate, Sm: Smooth, Sp: Sparse, T: Type, Up: Unicellular pointed, UT: Unicellular tabular, W: Wall and Wv: Wavy | |||||||||
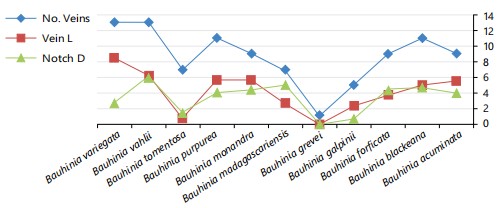
Vein patterns on the basal leaf surface, observed under a stereomicroscope, were rectinervous, curvinervous, or nervosous. The number of primary veins differs significantly among species. In bifoliate taxa, each leaflet contains a single primary vein, while in simple leaves, the number ranges from 0 to 15 (Fig. 2). The midrib length varies according to the depth of the apical notch and overall leaf length (Fig. 2). Leaf venation is generally palmiform, displaying either eucamptodromous or brochidodromous patterns. An exception is seen in B. grevei, where the venation is rectinervous; one leaflet is eucamptodromous, while the other is either uninervous-brochidodromous or trinervous-eucamptodromous.

|
| Table 3: | Leaf micro-morphological characters of the studied species | |||
| Ep cell | Hair | Stomata | ||||||||||
| Taxa | SH | PW | AW | OR | P | D | T | W | T | P | D | I |
| Bauhinia acuminata | Iso | Cv | St | Gr | Al | D | MUP | Sm | Pa | S | 115.8 | 11.8 |
| Bauhinia blakeana | Iso | Cv | St | Gr | Al | D | MUP | Sc | Pa and An | S | 182.7 | 18.4 |
| Bauhinia forficata | Iso | Fl | St | Gr | Al | D | MUP and Gl | Sm | Pa and An | S | 200.2 | 18.9 |
| Bauhinia galpinii | El | Cv | St | Sm | M | Sp | MUT | Sm | An | Su | 171.8 | 16.2 |
| Bauhinia grevei | Iso | Cv | St | Gr | M | Sp | Up | Gr | Ao | Su | 164.8 | 14.9 |
| Bauhinia madagascariensis | El | Cv | Si | Sm | Al | Mo | Gl | Sm | Ao | S | 209.8 | 19.2 |
| Bauhinia monandra | El | Fl | Wy | Gr | Al | D | UT | Gr | An | S | 228.7 | 21.2 |
| Bauhinia purpurea | Iso | Cv | Si | Gr | Al | Mo | Up | Sc | Pa and An | S | 195.8 | 19.2 |
| Bauhinia tomentosa | El | Cv | Wy | Gr | Al | D | Up | Gr | An | S | 192.2 | 18.5 |
| Bauhinia vahlli | Iso | Fl | Wv | Sm | Al | D | MUP and Gl | Sm | Ao | S | 171.6 | 17.6 |
| Bauhinia variegata | Iso | Cv | Si | Sm | Al | D | Up | Sm | An | S | 171.8 | 18.9 |
| Al: Allover, An: Anisocytic, Ao: Anomocytic, AW: Anticlinal wall, Cv: Convex, D: Dense, Fl: Flate, D: Density, Ep: Epidermal, Gl: Globular, Gr: Granulate, M: Margin, Mo: Moderate, MUP: Multicellular uniseriate glandular, MUT: Multicellular uniserriate tabular, P: Position, Pa: Paracytic, PW: Periclinal wall, S: Superficial, Sc: Scally, Si: Sinuate, Sm: Smooth, Sp: Sparse, T: Type, Up: Unicellular pointed, UT: Unicellular tabular, W: Wall and Wv: Wavy | ||||||||||||
| Key for the studied taxa according to their leaf characters |
| I-Spiny stipules | B. formica |
| I-Leafy stipules | |
| II-Bifoliate leaves | B. grevei |
| II-Simple leaves | |
| III-Number of main midribs in each leaflet 5 | B. galpinii |
| III-Number of main midribs in each leaflet 7 | |
| IV-Leaves small, their lengths never exceed 2.5 cm | B. tomentosus |
| IV-Leaves big, their lengths exceed 7 cm | B. madagascariensis |
| III-Number of main midribs 9 | |
| V-Leaf venation Eureticulodromous | B. acuminata |
| V-Leaf venation Brochidodromous | B. monandra |
| III-Number of main midribs 11 | |
| IV-Leaves big, their lengths more than 8.0 cm | |
| V-Leaf apex shallow notched 1.0-2.5 cm | B. variegata |
| V-Leaf apex deep notched wide V-shaped more than 3.5 cm | B. Blakeana |
| V-Leaf apex deep notched Y-shaped more than 3.5 cm sec | B. purpurea |
| III-Number of main midribs more than 11 | B. vahlii |
The leaf blade apices show varying degrees of V-shaped notches, ranging from narrow to very wide openings (Table 2, Fig. 3a-k).
Micro-morphology
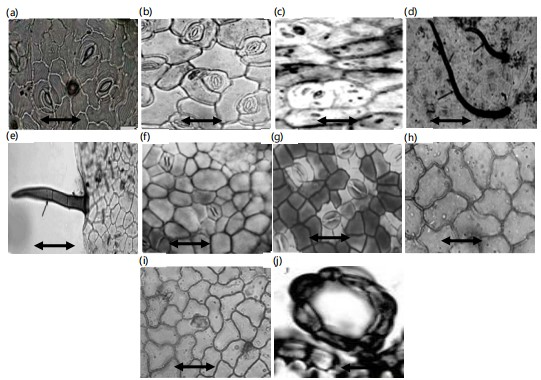
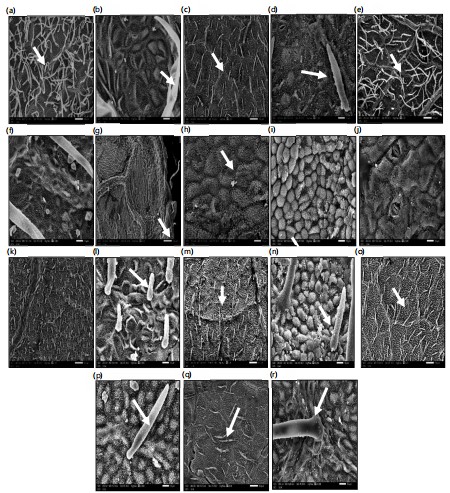
Epidermal cells: Photomicrographs obtained using both light microscopy and Scanning Electron Microscopy (SEM) of the studied Bauhinia leaf surfaces revealed that the epidermal cells are either elongated or isodiametric, and may be polygonal, tetragonal, or triangular, with straight, wavy, or undulate anticlinal walls (Fig. 4a-j). The periclinal walls are predominantly convex, as observed in SEM examinations (Table 3; Fig. 5b, h, i). However, in a few species B. forficata, B. monandra, and B. vahlii the periclinal walls appear flat (Fig. 5d, j) Secondary ornamentations on the periclinal walls were not visible, likely due to the presence of dense trichomes or waxy deposits. In the SEM images, convex epidermal cells appear globular in B. grevei and B. purpurea (Fig. 5i, j, o, p).
|
Trichomes: All the studied taxa exhibited varying densities of trichomes (hairs) on both the adaxial and abaxial leaf surfaces (Table 3). Trichomes were distributed across the entire leaf blade in most species; however, in B. galpinii and B. grevei, they were restricted to the leaf blade margins only (Fig. 5g). Hair density was classified as dense in B. acuminata, B. blakeana, B. forficata, B. monandra, B. tomentosa, B. vahlii, and B. variegata (Fig. 5a, c, e, k, m, o, q). In B. madagascariensis and B. purpurea, the trichome density was moderate, whereas in B. galpinii and B. grevei, it was sparse.
Each species typically exhibited a single type of trichome, except in B. forficata and B. vahlii, where two types were observed. The trichome types included uniseriate multicellular pointed, tabular, glandular, or unicellular pointed hairs (Fig. 2d, e). Cavitated trichomes were observed in B. forficata, B. madagascariensis, and B. vahlii (Fig. 2f). The walls of the trichomes varied: Granulate walls were recorded in B. grevei, B. monandra, and B. tomentosa (Fig. 5n), while scale-like trichomes were observed in B. purpurea (Fig. 5p).
Stomata: The leaf surfaces of the studied Bauhinia species are largely covered with epicuticular wax secretions, which obscure the visibility of stomata under scanning electron microscopy (SEM). Therefore, stomatal features were primarily examined using light microscopy. Photomicrographs obtained from both light and SEM revealed the presence of three stomatal types: Paracytic, anisocytic, and anomocytic. All species were hypostomatic, except B. blakeana, which exhibited amphistomatic distribution.
|
Stomata were randomly arranged and often obscured by dense trichomes in B. acuminata, B. blakeana, B. forficata, B. galpinii, B. monandra, B. vahlii, and B. variegata (Table 3). Some species B. blakeana, B. forficata, and B. purpurea possessed two stomatal types: Paracytic and anisocytic. In paracytic stomata, the two guard cells are flanked by two parallel subsidiary cells (Fig. 2c). In anisocytic stomata, the guard cells are surrounded by three subsidiary cells (Fig. 2b, i). Anomocytic stomata were recorded in B. grevei, B. madagascariensis, and B. vahlii, with four surrounding epidermal cells and no distinct subsidiary cells (Fig. 2g, j).
The stomata were generally small and located at the same level as the surrounding epidermal cells, except in B. galpinii and B. grevei, where the guard cells were sunken. Guard cells were consistently small in all taxa examined. The stomatal index was relatively low, and stomatal density, calculated as the number of stomata per 0.1 mm2, was also limited (Table 3). In all studied taxa, stomata appeared nearly closed, with very narrow apertures.
Morphological results according to SEM examination: Detailed leaf characteristics based on Scanning Electron Microscope (SEM) examination are summarized in Table 3 and illustrated in Fig. 5a-r. The epidermal cells are isodiametric or slightly elongated in all studied taxa, except in B. madagascariensis, where they exhibit a triangular outline (Fig. 5l). The periclinal walls are generally grooved, straight, or sinuate in most taxa, except in B. forficata, B. monandra, and B. vahlii, where they are superficial and straight (Fig. 5f, h). The anticlinal walls are typically convex; however, in taxa with superficial periclinal walls, such as B. forficata, the anticlinal walls appear flat (Fig. 5d).
Secondary ornamentations on the anticlinal walls vary across taxa. They are striate in B. galpinii (Fig. 5h), echinate in B. monandra (Fig. 5n), and smooth or granulate in the remaining species. Tertiary ornamentations are in the form of epicuticular wax secretions, which differ in shape and density or may be absent, as in B. galpinii, B. grevei, and B. monandra (Fig. 5h, j, n). These waxes appear as needle-like, flake-like, globular, or rosette/star-shaped projections (Fig. 5f, h, j).
Both adaxial and abaxial surfaces of the leaves are hairy in most of the studied taxa. Hairs are distributed across the entire leaf blade in most species; however, in B. galpinii and B. grevei, hairs are restricted to the leaf margins. Hair types are generally homogeneous, except in B. forficata and B. vahlii, where two types are observed multicellular uniseriate pointed hairs and globular hairs. Hair density and type vary significantly among taxa (Table 3). Multicellular uniseriate pointed hairs are long, soft, and woolly in appearance, whereas unicellular pointed or tabular hairs occur in denser trichome-covered species. In B. galpinii, a few multicellular uniseriate glandular hairs are restricted to the margins.
Globular unicellular hairs were observed on the leaf blades of B. madagascariensis and B. vahlii (Fig. 5o). The basal cells of the hairs are predominantly unicellular in most species, except in B. galpinii, where they are multicellular (Fig. 5n, p, r). Hair walls exhibit varying ornamentation patterns either psilate (smooth), granulate, or scaly.
DISCUSSION
The most important morphological characters in the identification of the taxa are the leaf. Leaf morphology lies in the first steps in the identification keys of the genera and species in many Floras. Meanwhile, leaf morphology indicates the degree of pollution and the input of environmental stress on the plant19,29. Many works have indicated the importance of studying leaf morphological variations as they have a direct effect on the plant physiological activities15,18. Not only are the leaf shapes and sizes affected by the surrounding environment, but also the leaf texture, density of hairs, stomatal density, and wax depositions are changed in response to the environmental disorders. Accordingly, the study of both the macro and micro-morphological characters is considered essential to evaluate the adaptive strategies of the plant to conquer the surrounding environment30. In this respect, this work has been done to use leaf macro and micro-morphological characters in the identification of the studied species and to evaluate both the adaptive degree to the polluted environment and the degree of evolution between the studied species.
The results obtained from this study revealed great variations in the leaf blade shape, sizes, texture, number of veins, and leaf apex notch. All the taxa have simple leaf with different sizes and surface texture. The leaf apices were notched with varied depth, except in B. grevei which has bilobed leaves with acute or rounded apices. The number of main veins, the length of the midrib and type of venation are useful in the differentiation of the studied species. Albert and Sharma31 pointed to the importance of both the number of leaf veins and the type of venation in the Bauhinia species in the identification of the taxa. They found that the number of main veins considered good character in the variation between the Bauhinia trees. These macro-morphological leaf variations are good characters in the differentiation between the studied species as seen in the identification key. According to the number of veins in the leaf blade, five groups can be recognized; the first group, with 0 to 5 veins and includes B. grevei and B. galpinii. The second group has leaf blades with 7 veins and has B. madgascariensis and B. tomentosa, while the third group has 9 veins and has B. acuminata and B. monandra. The last two groups have 11 veins and gathered B. blakeana, B. purpurea, and B. variegata, while the fifth group has more than 11 veins in the leaf blade and has both B. forficata and B. vahlii. This result was in partial agreement with Wunderlin5,6,9, as he divided the Bauhinia species into five sections.
The study of leaf characters is an important tool in understanding the degree of environmental pollution. Taia et al.19 found that the leaves underwent significant reduction in their measured length, width, area, and number of lateral veins compared with non-polluted sites. Uka and Belford29 found a reduction in the leaf area, stomata size, number, and index at the polluted areas. Meanwhile, they reported an increase in the number of epidermal and trichome lengths at the polluted areas. They considered these variations as pointers of environmental stress and could be used as indicators. From this work, the great variations in the leaf blade shape, sizes, texture, number of veins, and leaf apex notch can be considered as indications of the adaptation ways in these trees to adapt to the surrounding environment. Su et al.18 found that the leaf morphological traits can be considered as indications to their nutrient and physiological activities. Meanwhile Wang et al.32 correlated the density and number of stomata in both the abaxial and adaxial surfaces as indicator to the physiological activity in twelve Bauhinia species and three varieties. They used these stomatal indices in grouping these species into four groups, which agrees with this study.
The micro-morphological characters obtained from the SEM examination revealed that these characters are variable, especially hair density, basal cells, type of hairs, and hair wall ornamentations. Variations in the epidermal cells, periclinal and anticlinal walls, were limited in this study. Epicuticular wax depositions over the leaf surfaces varied between the taxa. Despite all of these micro-characters, they play of limited role in the division of the species. Hair density, type, and wall are diagnostic in a few species, especially the presence of globular cavitated secretory hairs, which were mentioned before by Marinho et al.33.
From the results obtained from this investigation, we can trace the adaptation level of the studied species. Yao-Qi and Zhi-Heng15 and Su et al.18 mentioned that the large-sized leaf blades are more adaptive to environmental pollution than the small leaves. As well, the many-nerved venation is considered an adaptive feature within the plants. Meanwhile, the hard, scaly leaf surface with dense hairs is considered from the plant’s strategy to protect it from the surrounding polluted environment. This study can give an idea about the evolutionary line within the studied taxa. Tucker14 pointed to the importance of using characters within Bauhinia species in tracing the evolutionary line between them. They advise considering genus Bauhinia as a natural group and undergone a reticulate pattern of variation. While8, considered genus Bauhinia as a paraphyletic one with the monospecific genus Brenierea, which is considered a sister to Bauhinia s.l. but they did not trace the evolutionary line within the Bauhinia species.
From the obvious leaf macro-morphological characters, we can postulate that the simple leaf blades with shallow apical notch or rounded apex, with palmate nerves as the primitive species, while the deepest apical notch or bifoliate leaves with reticulate nerves can be as more advanced.
CONCLUSION
The data obtained from this study supports the previous taxonomical studies of considering genus Bauhinia as a large genus with its division into two subgenera and five sections. Species within the genus show an evolutionary line as the simple leaves with a shallow apical notch, considered from the most primitive species. The notched apices with different depths are steps in the evolution toward the bifoliate leaves in B. grevei, which is considered the most advanced species within the studied taxa. The adaptive traits to the polluted environment in big cities are based on the large size of leaf blades with numerous main veins, scarious leaf texture, and densely hairy leaf surfaces with small guard cells.
SIGNIFICANCE STATEMENT
This study identified key leaf morphological traits that are diagnostic in differentiating Bauhinia taxa, which could be beneficial for improving species identification and enhancing the accuracy of traditional medicine applications. This study will assist researchers in uncovering critical areas of Bauhinia taxonomy, ecological adaptation, and environmental tolerance that have remained unexplored by many. Consequently, a new theory on the evolutionary adaptation of Bauhinia species to polluted environments may be developed.
REFERENCES
- Kumar, T. and K.S. Chandrashekar, 2011. Bauhinia purpurea Linn.: A review of its ethnobotany, phytochemical and pharmacological profile. Res. J. Med. Plant, 5: 420-431.
- Azani, N., M. Babineau, C.D. Bailey, H. Banks and A.R. Barbosa et al., 2017. A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny: The Legume Phylogeny Working Group (LPWG). TAXON, 66: 44-77.
- Larsen, K. and S.S. Larsen, 1991. Notes on the genus Bauhinia (Leguminosae-Caesalpinioideae) in SE Asia. Nord. J. Bot., 11: 629-634.
- Wang, Q., Z. Song, Y. Chen, S. Shen and Z. Li, 2014. Leaves and fruits of Bauhinia (Leguminosae, Caesalpinioideae, Cercideae) from the Oligocene Ningming Formation of Guangxi, South China and their biogeographic implications. BMC E Biol., 14.
- Wunderlin, R.P., 2009. Revision of Bauhinia subgenus Bauhinia section pauletia series araiaria (Cercideae: Caesalpinioideae: Fabaceae). J. Bot. Res. Inst. Texas, 3: 629-637.
- Wunderlin, R.P. and R.M. Eilers, 2009. Revision of Bauhinia subgenus Phanera section Schnella (Cercideae: Caesalpinioideae: Fabaceae). J. Bot. Res. Inst. Texas, 3: 619-628.
- Sinou, C., F. Forest, G.P. Lewis and A. Bruneau, 2009. The genus Bauhinia s.l. (Leguminosae): A phylogeny based on the plastid trnL-trnF region. Botany, 87: 947-960.
- Duarte-Almeida, J.M., G. Negri and A. Salatino, 2004. Volatile oils in leaves of Bauhinia (Fabaceae Caesalpinioideae). Biochem. Syst. Ecol., 32: 747-753.
- Wunderlin, R.P., 2010. Reorganization of the cercideae (Fabaceae: Caesalpinioideae). Phytoneuron, 48: 1-5.
- Sinou, C., W. Cardinal-McTeague and A. Bruneau, 2020. Testing generic limits in Cercidoideae (Leguminosae): Insights from plastid and duplicated nuclear gene sequences. TAXON, 69: 67-86.
- Igbari, A.D., W.O. Madu and O.T. Ogundipe, 2023. Systematic studies on some West African species of the tribe Bauhinieae (Cercidioideae). Webbia: J. Plant Taxon. Geogr., 78: 93-105.
- Trethowan, L., R. Clark and B. Mackinder, 2018. A synopsis of the neotropical genus Schnella (Cercideae: Caesalpinioideae: Leguminosae) including 12 new combinations. Phytotaxa, 204: 237-252.
- Meng, H.H., F.M.B. Jacques, T. Su, Y.J. Huang, S.T. Zhang, H.J. Ma and Z.K. Zhou, 2014. New biogeographic insight into Bauhinia s.l. (Leguminosae): Integration from fossil records and molecular analyses. BMC E Biol., 14.
- Tucker, S.C., 2002. Floral ontogeny of Cercis (Leguminosae: Caesalpinioideae: Cercideae): Does it show convergence with papilionoids? Int. J. Plant Sci., 163: 75-87.
- Yao-Qi, L. and W. Zhi-Heng, 2021. Leaf morphological traits: Ecological function, geographic distribution and drivers. Chin. J. Plant Ecol., 45: 1154-1172.
- Hickey, L.J., 1973. Classification of the architecture of dicotyledonous leaves. Am. J. Bot., 60: 17-33.
- Fritsch, P.W., C.F. Nowell, L.S.T. Leatherman, W. Gong, B.C. Cruz, D.O. Burge and A. Delgado-Salinas, 2018. Leaf adaptations and species boundaries in North American Cercis: Implications for the evolution of dry floras. Am. J. Bot., 105: 1577-1594.
- Su, Y., M. Renz, B. Cui, X. Sun, Z. Ouyang and X. Wang, 2021. Leaf morphological and nutrient traits of common woody plants change along the urban-rural gradient in Beijing, China. Front. Plant Sci., 12.
- Taia, W.K., W.M. Amer, A.M. Abd El-Maged and A.B. Hamed, 2024. Comparative study on the effect of pollution on the leaf morphology of common ornamental species in both Giza District, Cairo and Smouha, Alexandria, Egypt. Taeckholmia, 44: 48-72.
- Smýkal, P., C.J. Coyne, M.J. Ambrose, N. Maxted and H. Schaefer et al., 2015. Legume crops phylogeny and genetic diversity for science and breeding. Crit. Rev. Plant Sci., 34: 43-104.
- Feodorova, T.A. and O.S. Alexandrov, 2020. Comparative studying of leaf trichomes, teeth and glands in Populus nigra L., Populus deltoides W. Bartram ex Marshall and their hybrids. Forests, 11.
- El-Gazzar, A. and N.A. Moustafa, 2021. The taxonomic significance of gross morphology, indumentum and cellular inclusions in Solanum L. (Solanaceae). Egypt. J. Bot., 61: 759-771.
- Taia, W.K. and G.E. El-Badan, 2023. Macro-morphological leaf variations between Bombacaceae and Malvaceae with their significance in taxonomy and phylogeny. Egypt. Acad. J. Biol. Sci. C Physiol. Mol. Biol., 15: 923-932.
- Taia, W.K., A. Hassan and G.E. El-Badan 2023. Leaf variations within representative genera of tribe Malveae and their significance in the taxa phylogeny. Egypt. J. Bot., 63: 727-742.
- Taia, W.K., A.A. Shehata and E.M. Bassiouni, 2023. Micromorphological investigation in leaf epidermis of some monocotyledons taxa. Egypt. J. Bot., 63: 951-968.
- Uphof, J.C.T., 1962. Plant Hairs. Schweizerbart Science, Berlin, Germany, ISBN: 978-3-443-39005-1, Pages: 292.
- Dilcher, D.L., 1974. Approaches to the identification of angiosperm leaf remains. Bot. Rev., 40: 1-157.
- Punt, W., P.P. Hoen, S. Blackmore, S. Nilsson and A. le Thomas, 2007. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol., 143: 1-81.
- Uka, U.N. and E.J.D. Belford, 2020. Response of roadside tree leaves in a tropical city to automobile pollution. Notulae Sci. Biol., 12: 752-768.
- Prigioniero, A., A. Postiglione, D. Zuzolo, Ü. Niinemets and M. Tartaglia et al., 2023. Leaf surface functional traits influence particulate matter and polycyclic aromatic hydrocarbons air pollution mitigation: Insights from Mediterranean urban forests. J. Cleaner Prod., 418.
- Albert, S. and B. Sharma, 2013. Comparative foliar micromorphological studies of some Bauhinia (Legiminoseae) species. Turk. J. Bot., 37: 276-281.
- Wang, Y.H., S. Wicke, H. Wang, J.J. Jin and S.Y. Chen et al., 2018. Plastid genome evolution in the early-diverging legume subfamily Cercidoideae (Fabaceae). Front. Plant Sci., 9.
- Marinho, C.R., R.B. Oliveira and S.P. Teixeira, 2016. The uncommon cavitated secretory trichomes in Bauhinia s.s. (Fabaceae): The same roles in different organs. Bot. J. Linn. Soc., 180: 104-122.
How to Cite this paper?
APA-7 Style
Taia,
W.K., Bassioni,
E.M. (2025). Leaf Morphological Variations Among Selected Bauhinia L. Species. Research Journal of Botany, 20(1), 180-190. https://doi.org/10.3923/rjb.2025.180.190
ACS Style
Taia,
W.K.; Bassioni,
E.M. Leaf Morphological Variations Among Selected Bauhinia L. Species. Res. J. Bot 2025, 20, 180-190. https://doi.org/10.3923/rjb.2025.180.190
AMA Style
Taia
WK, Bassioni
EM. Leaf Morphological Variations Among Selected Bauhinia L. Species. Research Journal of Botany. 2025; 20(1): 180-190. https://doi.org/10.3923/rjb.2025.180.190
Chicago/Turabian Style
Taia, Wafaa, Kamal, and Eman Mohammed Bassioni.
2025. "Leaf Morphological Variations Among Selected Bauhinia L. Species" Research Journal of Botany 20, no. 1: 180-190. https://doi.org/10.3923/rjb.2025.180.190

This work is licensed under a Creative Commons Attribution 4.0 International License.