Infectivity of the Fusarium fujikuroi Species Complex (FFSC) on Maize (Zea mays Lam.) in Gauteng, South Africa
-
Yakubu Pwajok Mwanja


Department of Microbiology, Faculty of Natural and Applied Sciences, Plateau State University, Bokkos, Nigeria
Vou Moses ShuttDepartment of Plant Science and Biotechnology, Faculty of Natural Science, University of Jos, Jos, Nigeria
Filibus MusaDepartment of Microbiology, Faculty of Natural and Applied Sciences, Plateau State University, Bokkos, Nigeria
Philomena Daniel AyikaDepartment of Microbiology, Faculty of Natural and Applied Sciences, Plateau State University, Bokkos, Nigeria
| Received 30 Jul, 2024 |
Accepted 10 Jan, 2025 |
Published 31 Jan, 2025 |
Background and Objective: Of the world’s top ten phytopathogenic fungi, Fusarium is the only group represented by more than one genus causing yield losses on crops and also producing a wide range of mycotoxins in food and feed products which can cause deleterious effects on human and livestock. The study aimed to investigate the pathogenicity of some Fusarium species isolated from Gauteng, South Africa on maize. Materials and Methods: A complete randomized block design with 26 isolates, belonging to the members of the Fusarium fujikuroi species complex (FFSC) was screened in 2017 for their infectivity on maize in Plant Protection Research Institute (PPRI), Pretoria, South Africa. The parameters tested included germination efficiency, plant height, dry weight, and disease severity. The results were analyzed using Duncan’s Multiple-Range Test at a 5% level of significance. Results: The infection with F. burgessii and F. nygamai isolates PPRI 21048 and PPRI 21265 produced the highest and lowest disease severity index of 72.89% and 41.33%, respectively. These caused a reduction in plant height by 76.07% and 44.22%, as well as a reduction in dry weight by 58.24% and 20.50%. There was a significant difference (p<0.05) amongst the species and isolates of the same species. A low, yet significant negative correlation was observed both between disease severity and plant height (r = -0.6463, p = 0.0004) on the one hand and between disease severity and dry weight (r = -0.5038, p = 0.0087) on the other hand. Conclusion: The Fusarium species have caused varying degrees of infectivity on maize from a reduction in plant height to a reduction in dry weight with significant differences among the Fusarium species and isolates of the same species.
| Copyright © 2025 Mwanja et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
The fungal genus Fusarium is one of the most economically important fungal genera because of the yield losses it causes on crops due to its plant pathogenic activity. Of the top ten plant-pathogenic fungi, Fusarium has two spots while the other eight have one spot each, thus making it one of the most important fungi affecting a huge diversity of crops worldwide1. Fusarium produces a wide range of mycotoxins in food and feed products which could cause fatal effects in humans and livestock2. Two Fusarium species namely F. graminearum, which is the causative agent of head blight of wheat, and F. oxysporum, which is the causative agent of a range of wilt and stem rot diseases have been listed in the world’s top ten most important plant-pathogenic fungi3. The American Phytopathology Society’s list of diseases of agricultural and horticultural crops shows that 83 of 108 plant species representing 76.85% of the plants surveyed have one or more Fusarium diseases affecting their production. Fusarium species produce a range of mycotoxins such as trichothecene and fumonisin notably4. The former is a protein synthesis inhibitor while the latter causes fatal livestock diseases and is considered potentially carcinogenic for humans2,5. Some Fusarium species also cause diseases with devastating effects in humans especially those who are immunocompromised6.
Because of the foregoing, it is critically important for Fusarium to have a taxonomy that is stable for easy identification and diagnosis of diseases. This will lead to a proper management of the diseases caused by Fusarium species with a greater percentage of confidence. Unfortunately, this has not been the case because of the confusing taxonomy and systematics of Fusarium. For example, the genus has witnessed a decline in the number of species in the years between the 1920s and 1950s from a little more than a thousand species to nine species7. The application of multi-locus molecular phylogenetics in the past twenty years has revolutionized the studies of Fusarium species diversity, evolutionary relationships, and genealogical concordance phylogenetic species recognition8, with only half of the estimated 300 genealogically exclusive phylogenetic species formally described7,9.
The role of mycotoxins in plant-pathogen interaction is unclear but some researchers have shown that it plays a role to the benefit of the pathogen enabling it to progress in plant tissues10,11. This was demonstrated by Harris et al.11 and Proctor et al.12 in their work on trichothecene–producing fungi in which they showed that some F. graminearum strains caused more disease in maize than their trichothecene non-producing mutants that were pathogenic. Although Adams and Hart13 have reported that deoxynivalenol (DON) was not a pathogenicity and virulence factor for F. graminearum in maize, Mesterhazy10 has reported that there was a significant correlation between the virulence of F. graminearum and F. culmorum and their DON and nivalenol (NIV) content in wheat grain. The role of fumonisin in pathogenicity, phytotoxicity, and virulence is unclear even though it is phytotoxic in maize seedlings14,15. These researchers have further reported the inhibitory effect of fumonisin on root hair development and root growth. In contrast, Van-Asch et al.16 have reported the phytotoxic effect of fumonisin on maize callus in culture with further phytotoxicity symptoms on seedlings being induced when the seedlings were watered with high concentrations of the mycotoxin even in the absence of the pathogen15. Fumonisin production by F. verticillioides is a necessity for the development of foliar disease symptoms in maize seedlings17 but may not be essential for pathogenicity to maize seedlings as argued by Desjardins et al.18 Further studies by these researchers have shown that the progeny of a fum1 positive field strain of F. verticillioides with high levels of virulence were found to be associated with fumonisin production. Still, no progeny of a fum1 negative field strain of F. verticillioides were observed to have high levels of virulence. The study aimed to investigate the pathogenicity of some Fusarium species isolated from the grassland biome of South Africa on maize.
MATERIALS AND METHODS
Study area: The study was conducted at the University of Johannesburg, South Africa between January, 2017 and December, 2020.
Fungal isolates and plant material: A collection of 26 Fusarium fujikuroi species complex (FFSC) isolates used in this study was sourced from the Plant Protection Research Institute (PPRI) of the Agricultural Research Council (ARC), Pretoria, South Africa. All the isolates F. nygamai (12), F. bacteroides (10), and F. burgessii (4) originated from the soils of the Willem Pretorius Nature Reserve, Free State Province, South Africa (Table 1). The isolates were tested in greenhouse trials for pathogenicity on commercial seeds of maize (cultivar “PAN 8816").
| Table 1: | Fusarium strains analyzed in this study indicate species and strain | |||
| Fusarium species | Strainac |
| Fusarium nygamai | PPRI 19184ac |
| Fusarium nygamai | PPRI 19196ac |
| Fusarium nygamai | PPRI 19219ac |
| Fusarium nygamai | PPRI 20533ac |
| Fusarium nygamai | PPRI 20740ac |
| Fusarium nygamai | PPRI 21056ac |
| Fusarium nygamai | PPRI 21098ac |
| Fusarium nygamai | PPRI 21255ac |
| Fusarium nygamai | PPRI 21265ac |
| Fusarium nygamai | PPRI 21294ac |
| Fusarium nygamai | PPRI 21308ac |
| Fusarium nygamai | PPRI 21573ac |
| Fusarium bacteroides | PPRI 19149ac |
| Fusarium bacteroides | PPRI 19231ac |
| Fusarium bacteroides | PPRI 21033ac |
| Fusarium bacteroides | PPRI 21055ac |
| Fusarium bacteroides | PPRI 21071ac |
| Fusarium bacteroides | PPRI 21253ac |
| Fusarium bacteroides | PPRI 21272ac |
| Fusarium bacteroides | PPRI 21300ac |
| Fusarium bacteroides | PPRI 21306ac |
| Fusarium bacteroides | PPRI 22756ac |
| Fusarium burgessii | PPRI 21048ac |
| Fusarium burgessii | PPRI 21079ac |
| Fusarium burgessii | PPRI 21097ac |
| Fusarium burgessii | PPRI 21263ac |
| a: Plant Protection Research Institute (PPRI), Agricultural Research Council (RC), Pretoria, South Africa and c: Originated from willem pretorius nature reserve, free state, South Africa | |
Inoculum preparation: The method of Nel et al.19 was used to prepare the inoculum with slight modifications. The Fusarium isolates germinated from a single macroconidium were sub-cultured and grown on PDA under cool-white and near-ultra violet fluorescent lights at 25±°C for 7 days, after which the mycelia and conidia were harvested from the cultures by adding 20 mL of sterile distilled water and gently scraping the surface with a sterile Drigalski spatula into a sterile 100 mL beaker. The harvested mycelia and conidia were used to inoculate 250 g sterilized millet seeds in a 500 mL sterile Erlenmeyer flask. The flask was covered with cotton wool, wrapped with foil paper, and then incubated in the dark at 24°C for 24 days. The infested millet seeds were ground to powder using a blender (Salton 450W, Model: SB400E) and added to steam-sterilized soil at the rate of 22 g 1000/g soil. The inoculated soil was mixed and dispensed in 12.5 cm diameter plastic pots to produce a 2.2% inoculum concentration in the soil. The soil consisted of a mixture of potting and compost soil in a ratio of 1:1.
In vivo pathogenicity assay: The greenhouse trials were carried out using three biological repeats (bioreps) with 2 week intervals between them. The trials consisted of three inoculated 12.5 cm plastic pots per isolate and each pot was planted with 5 seeds and was watered three times a week for 4 weeks. The experiments were laid out in a complete randomized design consisting of 26 treatments and three replications. The control consisted of three 12.5 cm plastic pots dispensed with ground un-inoculated sterilized millet seeds mixed with steam-sterilized soil at a 2.2% inoculum concentration in the soil.
Disease symptoms were visually assessed and rated on a scale of 0-52, with slight modifications in which 5 = Dead seedlings; 4 = 76-100% roots showing symptoms, and 3.1->5 cm necrotic lesion size on leaf; 3 = 51-75% roots showing symptoms and 1.1-3.0 cm necrotic lesion size on leaf; 2 = 26-50% roots showing symptoms and 0.1-1.0 cm necrotic lesion size on leaf; 1 = 1-25% roots showing
symptoms; 0 = healthy plant with no symptoms. Disease severity, seed germination, plant height as well as dry weight were measured. Data were converted to a disease severity index score using the formula reported by Liu et al.20. The disease severity index was calculated from the rating by the following formula:
Microscopy
Killing and fixation: The Glycol Methacrylate (GMA) method21 was used to prepare the root samples. Ten root samples were collected from the greenhouse at the end of the pathogenicity trials. Root pieces 2-4 cm long were cut and surface sterilized using 0.1% Sodium Hypochlorite (NaOCl), and rinsed three times with distilled water. The samples were then fixed in FAA fixative solution (10 mL of 40% formaldehyde, 5 mL of 99% glacial acetic acid, 50 mL of 99.9% ethanol, and 35 mL of sterile distilled water) for 48 hrs.
Dehydration: The samples were dehydrated by processing in a graded series of ethanol twice for 6 hrs in each of 70 and 100%. This was followed by processing in 100% n-propanol twice for 6 hrs and final processing in 100% n-butanol for 6 hrs.
Infiltration and embedding: The root samples were covered with GMA using the first infiltration GMA and placed in a dark cupboard for 24 hrs. The second and third infiltration GMA was used to replace the first infiltration GMA and placed in a dark cupboard for 24 hrs. The root samples were embedded in GMA in labeled gelatine capsules and placed in an oven (‘Brickovens’ - New York, USA) set at 60°C for 24 hrs.
Sectioning: Section (5 μm thick) was done using the microtome. Blocks were prepared in GMA in gelatine capsules and thin sections 5 μm thick were cut with the help of a microtome (SORVALL “PORTER-BLUM” Ultra Microtome MT-1 Connecticut, USA). At least 10 slides were prepared for each sample.
Staining and mounting: The slides were placed in saturated 2, 4-dinitrophenylhydrazine (DNPH) solution for 30 min and then washed 5 times in tap water. Slides were placed in 1% periodic acid solution for 10 min and washed 3 times in tap water. The slides were dissolved in Schiff’s reagent for 30 min and followed by 31 min washes with tap water. The slides were dried on the slide warmer, stained with 0.05% toluidine blue for 3 min, and followed by three 1 min washes with tap water until the water was more or less colorless. The slides were dried overnight on the slide warmer. After drying the slides, they were mounted with an entellan synthetic resin, covered with cover slips, and viewed on the microscope. Observations were made using an OLYMPUS microscope (CX41RF, Tokyo, Japan) coupled to a camera (OLYMPUS Stream Essentials 2.2.3 Build 17023).
Statistical analysis: Data from the pathogenicity assay were subjected to Analysis of Variance (ANOVA) using the program Minitab 16 (Minitab Inc. Pennsylvania, USA). Disease index percentages were arcsine-transformed before ANOVA. Mean values in each treatment were compared by the Least Significant Difference (LSD) and Duncan’s New Multiple Range (DNMR) test at a 5% (p = 0.05) level of significance. Pearson’s linear correlation coefficient was used to evaluate the relationship between the disease severity index and the disease parameters
RESULTS
All the 26 Fusarium fujikuroi species complex (FFSC) isolates under investigation in this study originated from the Willem Pretorius Nature Reserve (Table 1) and were able to infect maize and produce characteristic disease symptoms such as disease severity index (Fig. 1), correlation of disease severity with plant height (Fig. 2), zero as well as low germination, root rots (Fig. 3), tip and blade necrotic lesions (Fig. 4), reduced/stunted aerial and root growth (Fig. 5), and reduced plant biomass (dry weight). The levels of aggressiveness differed between species and between isolates of the same species.

|
|
In vivo pathogenicity assay
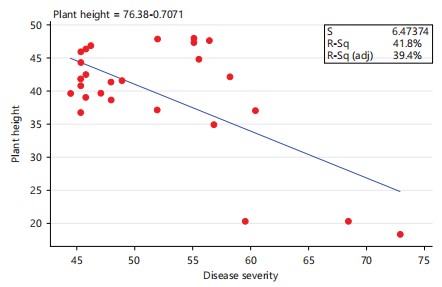
Disease severity: The highest maize disease severity among the three Fusarium species under investigation manifested by root rots symptoms (Fig. 3), as w ell as leaf tip and leaf blade necrotic lesions (Fig. 4) was observed in F. burgessii isolate PPRI 21048 (72.89%), followed by F. bacteroides and F. nygamai isolates PPRI 21306 and PPRI 21308 (68.44% and 61.33%), respectively (Fig. 1). These isolates were responsible for the r eduction in height of the plants (Fig. 5). The lowest disease severity among the three tested Fusarium species was observed in F. nygamai isolate PPRI 21265 (41.33%), followed by F. bacteroides isolate PPRI 21253 and F. burgessii isolate PPRI 21263 (42.22%) (Fig. 1). Further disease severity in maize included isolates PPRI 19196 of F. nygamai (56.11%); PPRI 19149 and PPRI 21033 of F. bacteroides (56.44% and 55.11%, respectively) and PPRI 21097 of F. burgessii (60.44%) (Fig. 1). There was a low, yet significant negative correlation between disease severity and plant height of maize (Fig. 2) (r = - 0.6463, p = 0.0004).
|

|
Plant height: The isolates caused a reduction in both aerial growth and root growth (Fig. 3-5). The highest plant height was observed in F. bacteroides isolate PPRI 21033, (48.0267 cm), followed by the same F. bacteroides isolate PPRI 19231 (47.9333 cm) in second position, while F. bacteroides isolates PPRI 19149 and PPRI 22756 (47.6767 cm and 47.44 cm) emerged in the third and fourth positions, respectively (Fig. 6). These isolates caused the least damage thereby producing the tallest maize hosts. The lowest maize plant height was observed in F. burgessii isolate PPRI 21048 (18.2523 cm), followed by F. bacteroides isolate PPRI 21306 (20.2733 cm) in the second lowest position while F. nygamai isolates PPRI 21308 (20.3633 cm) emerged in the third lowest position (Fig. 6). These isolates caused more damage thereby producing the shortest hosts (Fig. 6). Infection of maize with F. nygamai isolates PPRI 21308, PPRI 21056 and PPRI 20533 produced plant heights of 20.36 cm, 36.79 cm and 37.12 cm which represent 26.7%, 48.23% and 48.66%, respectively, when compared with the control (76.28 cm). Similarly, infection with F. bacteroides isolates PPRI 21306 and PPRI 21272 produced plant heights of 20.27 cm and 40.73 cm which represent 26.57% and 53.39%, respectively, while maize plants infected with F. burgessii PPRI 21048 and PPRI 21079 produced heights of 18.25 cm and 34.98 cm which represent 23.92% and 45.86%, respectively (Fig. 6). There was a low, yet significant negative correlation between maize plant height and disease severity (Fig. 2) (r = -0.6463, p = 0.0004).
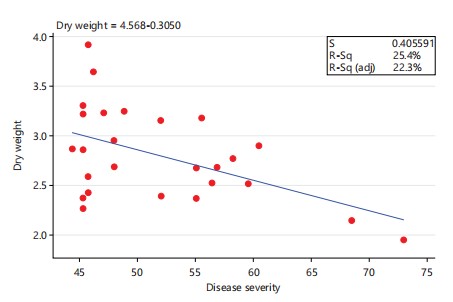
Dry weight: The highest dry weight among the three Fusarium species under investigation was observed in F. nygamai isolate PPRI 21265 (4.0837 g), followed by F. bacteroides isolate PPRI 21253 (3.819 g) in the second position while F. nygamai and F. burgessii isolate PPRI 19184 and PPRI 21263 (3.4823 and 3.4247 g) occupied the third and fourth positions, respectively (Fig. 7).

|

|
These isolates caused least damage to their maize plant hosts. The lowest maize dry weight was observed in F. burgessii isolate PPRI 21048 (2.145 g), followed by F. bacteroides isolates PPRI 21306 and PPRI 21071 (2.3337 g and 2.4537 g) in the second and third lowest positions, respectively (Fig. 7). These isolates caused more damage to their maize plant host. Infection of maize plants with Fusarium nygamai isolates PPRI 21308, PPRI 21255, PPRI 21056, PPRI 21573 and PPRI 19196 produced reduced dry weights of 2.706 g, 2.7733 g, 2.816 g, 2.871 g and 2.9557 g, which represent 52.40%, 53.71%, 54.53%, 55.60%, and 57.24%, respectively, when compared to the control (5.1637 g) (Fig. 7). Similarly, F. bacteroides isolates PPRI 21306 and PPRI 21071 infection of maize plant hosts produced reduced dry weights of 2.3337 g and 2.4536 g which represent 45.19% and 47.52%, respectively, whereas infection with F. burgessii isolates PPRI 21048 and PPRI 21079 resulted in a reduction in dry weights of 2.145 g and 2.8607 g, which represent 41.54% and 55.40%, respectively, when compared with the control. There was no significant difference (p>0.05) between isolates of the same species. A low, yet significant negative correlation was observed between disease severity and dry weight (Fig. 8) (r = -0.5038, p = 0.0087).

|
|
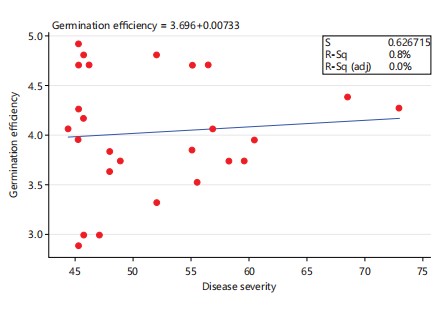
Germination efficiency: The highest maize seed germination efficiency among the three Fusarium species under investigation was observed in F. bacteroides isolates PPRI 21071 and PPRI 21033 (4.7778 and 4.6667) in the first and second positions, respectively, while four F. bacteroides isolates PPRI 19231, PPRI 21055, PPRI 21300 and PPRI 22756 tied in the third position with 4.4444 (Fig. 9). Infection of maize with F. nygamai isolates PPRI 20533, PPRI 21294 and PPRI 21573 produced reduced seed germination of 2.3333, 2.7778 and 3.1111, which represent 46.67%, 55.56% and 62.22%, respectively, when compared to the control (5.0000) (Fig. 9). Infection of maize with F. bacteroides isolates PPRI 21272 and PPRI 21253 produced reduced seed germination of 2.4444 and 4.0000, which represent 48.89% and 80%, while maize plants infected with F. burgessii isolates PPRI 21079 and 21263 produced seed germination of 3.1111 and 3.4444, which represent 62.22% and 68.89%, respectively (Fig. 9). There was no significant difference (p>0.05) between the isolates of the same species. A low, yet not significant positive correlation was observed between the maize germination efficiency and disease severity (Fig. 10) (r = 0.0904, p = 0.6606).

|
|
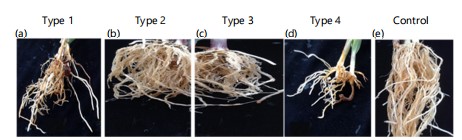
Microscopy: At 28 DAI the test isolates had demonstrated different levels of colonization of the different root tissues of the host. Fusarum nygamai hyphae localized colonization in the cells of the cortex only (Fig. 11a), whereas F. bacteroides has already invaded and digested most of the cells of the cortex creating gaps within the cortex (Fig. 11b). Fusarum bacteroides was restricted to the cortex and was not able to penetrate the endodermal layer to colonize the central cylinder (Fig. 11b). Fusarum burgessii at this stage had not only colonized and digested most of the cells of the cortex but had also penetrated the endodermal thick-walled layer and entered the central cylinder (Fig. 11c). Fusarum burgessii had massively destroyed the cortical cells and created gaps leaving the remnant of cells within the cortex (Fig. 11c). The maize control root tissues were intact and completely free of colonization by hyphae (Fig. 11d).

|
Fusarum bacteroides has caused digestion of the cells of the cortex leaving some gaps and remnants of undigested cells in the cortex (arrows) at 28 DAI. The central cylinder was also colonized by hyphae. Infection with F. burgessii caused massive digestion and degradation of the cells of the cortex leaving gaps and remnants of cortical cells in the cortex (arrows). The inner thick wall of the endodermal cells (E) was reduced to a thin layer. The cells of the central cylinder were also invaded (arrows).
Part of the control root is shown devoid of hyphae in the cortex (C), the endodermis (E), and the central cylinder which are all intact. The thick-walled endodermis (E) separates the cortex from the central cylinder. V, vessels; R, rhizodermis; a, b, c, and d are cross sections of maize roots infected with F. nygamai, F. bacteroides, F. burgessii, and the control, respectively. Scale bars = 50 μm.
DISCUSSION
Many Fusarium species have been associated with grain crops in South Africa22. Those that affect maize include F. verticillioides (Sacc.) Nirenberg; F. subglutinans (Wollenweber & Reinking) Nelson, Toussan and Marasas. Infection with F. burgessii (PPRI 21048) and F. nygamai (PPRI 21265) resulted in the highest (72.89%) and lowest (41.33%) disease severity indices, reducing plant height by 76.07% and 44.22% and dry weight by 58.24% and 20.50%, respectively. Significant differences (p<0.05) were observed among species and isolates, with a negative correlation between disease severity and both plant height (r = -0.6463, p = 0.0004) and dry weight (r = -0.5038, p = 0.0087).
The recovery of more isolates of F. nygamai (46.15%) than the isolates of F. bacteroides (38.46%) and F. burgessii (15.38%) in the roots coupled with the fact that F. nygamai limited its infection to the epidermal and cortical cells, in this study, refutes the report of Leyva-Madrigal et al.23 in which they showed the ability of F. nygamai to colonize the vascular tissues among two other Fusarium pathogens of cereal crops that only limited their infection to the epidermis and adjacent cortical cells.
The isolates of F. bacteroides and F. burgessii have only been associated with root rot and leaf tip and blade necrosis in maize in South Africa in this study. The plants infected with the two latter Fusarium species developed the same symptomatology as the plants infected with F. nygamai (root rot, leaf tip, and blade necrotic lesion, and reduced aerial and root growth) but at varying degrees of infectivity. In this study, the isolates PPRI 21048 of F. burgessii, PPRI 21306 of F. bacteroides and PPRI 21308 of F. nygamai which produced the highest disease severity also produced the lowest plant height and plant biomass (dry weight) which was consistent with the report of Leyva-Madrigal et al.23.
The mean seed germination showed that there was no significant difference (p>0.05) between species and among isolates of the same species (Fig. 9). No correlation was observed between the disease severity and seed germination (Fig. 10) (r = 0.0904, p = 0.6606), The low, yet significant negative correlation observed in this study between disease severity and plant height (Fig. 2) (r = -0.6463, P = 0.0004), was consistent with the report of Leyva-Madrigal et al.23 where they observed a low, significant negative correlation between plant height of maize and the disease severity in a mixed infection trials of maize with F. verticillioides, F. nygamai, F. thapsinum and F. andiyazi in Mexico.
Infection with F. burgessii isolate PPRI 21048 produced the most reduced dry weight (biomass). A significant negative correlation was observed between disease severity and dry weight (Fig. 8) (r = -0.5038, p = 0.0087). This refutes the report of Leyva-Madrigal et al.23 in which they observed no correlation between disease severity and shoot and root biomass in maize mixed infection trials. Fusarium nygamai infection was limited to the cortex, whereas F. bacteroides and F. burgessii had access to the central cylinder confirming previous observations of Elzein et al.24. Cell wall degrading enzymes (CWDEs) such as xylanases, and proteases and polygalacturonases are secreted by Fusarium species during the penetration and colonization of host roots25. The ability of F. nygamai to colonize and digest cortical cells but not the endodermal cells and the central cylinder could be attributed to the fact that the enzymes required were not expressed at the point of the infection cycle or that the enzymes required were absent as can be evidenced by the report of Ruiz-Roldan et al.26 where they demonstrated the differential expression of two genes xyl3 and xyl2 which encode xylanases in tomatoes infected with F. oxysporum f. sp. lycopersici. The former gene was expressed throughout the disease cycle while the latter gene was expressed only during the final stages of the disease cycle, which is an indication that genes are differentially expressed during a disease cycle.
To the best of our knowledge, this is the first report of the infectivity of these three Fusarium species in the grassland biome of South Africa. Pathogenicity of F. nygamai on sorghum in South Africa has been reported earlier22 but there is little or no accompanying data regarding the pathogenicity of F. bacteroides and F. burgessii on grain crops in South Africa22.
Fusarims burgessii has shown more aggressiveness followed by F. bacteroides and F. nygamai in that order in this study. The low aggressiveness of F. nygamai compared with its infectivity in the report of Beukes et al.22 could be attributed to the differences in host preferences27,28. Further pathogenicity trials on other grain crops are suggested to enable robust accompanying data on the pathogenicity of the members of the FFSC on grain crops in South Africa.
CONCLUSION
Fusarium species exhibited varying degrees of pathogenicity on maize in Gauteng, South Africa, causing significant reductions in plant height and dry weight. Among the species studied, F. burgessii caused the most severe disease. This is the first report of the pathogenicity of these Fusarium species on maize in this region, highlighting the need for further studies on their impact and pathogenicity on diverse grain crops. Such investigations are essential to address food security challenges and improve maize production in the area.
SIGNIFICANCE STATEMENT
This study highlights the significant impact of Fusarium species, particularly F. burgessii, on maize production in Gauteng, South Africa. These pathogens not only cause severe yield losses but also produce mycotoxins that threaten food safety and security. As the first report of these species' pathogenicity on maize in the region, it underscores the need for further research to develop effective management strategies and support sustainable maize production.
ACKNOWLEDGMENT
Special appreciation goes to the Plant Protection Research Institute (PPRI), Agricultural Research Council (ARC), Pretoria, South Africa for providing the Fusarium isolates for this research.
REFERENCES
- Dean, R., J.A.L. van Kan, Z.A. Pretorius, K.E. Hammond-Kosack and A. di Pietro et al., 2012. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol., 13: 414-430.
- Munkvold, G.P., 2017. Fusarium Species and their Associated Mycotoxins. In: Mycotoxigenic Fungi: Methods and Protocols, Moretti, A. and A. Susca (Eds.), Humana Press, New York, ISBN: 978-1-4939-6707-0, pp: 51-106.
- Marasas, W.F.O., P.E. Nelson and T.A. Toussoun, 1984. Toxigenic Fusarium Species: Identity and Mycotoxicology. Pennsylvania State University Press, University Park, Pennsylvania, ISBN: 9780271003481, Pages: 328.
- Desjardins, A.E., 2006. Fusarium Mycotoxins: Chemistry, Genetics, and Biology. APS Press, Saint Paul, Minnesota, ISBN: 9780890543351, Pages: 260.
- O’Donnell, K., A.P. Rooney, R.H. Proctor, D.W. Brown and S.P. McCormick et al., 2013. Phylogenetic analyses of RPB1 and RPB2 support a middle Cretaceous origin for a clade comprising all agriculturally and medically important fusaria. Fungal Genet. Biol., 52: 20-31.
- Summerell, B.A. and J.F. Leslie, 2011. Fifty years of Fusarium: How could nine species have ever been enough? Fungal Diversity, 50: 135-144.
- Aoki, T., K. O’Donnell and D.M. Geiser, 2014. Systematics of key phytopathogenic Fusarium species: Current status and future challenges. J. Gen. Plant Pathol., 80: 189-201.
- O’Donnell, K., S.P. McCormick, M. Busman, R.H. Proctor and T.J. Ward et al., 2018. Marasas et al. 1984 “Toxigenic Fusarium species: Identity and mycotoxicology” revisited. Mycologia, 110: 1058-1080.
- Wolf, J.C. and C.J. Mirocha, 1973. Regulation of sexual reproduction in Gibberella zeae (Fusarium roseum 'Graminearum') by F-2 (Zearalenone). Can. J. Microbiol., 19: 725-734.
- Mesterházy, Á., 2002. Role of deoxynivalenol in aggressiveness of Fusarium graminearum and F. culmorum and in resistance to Fusarium head blight. Eur. J. Plant Pathol., 108: 675-684.
- Harris, L.J., A.E. Desjardins, R.D. Plattner, P. Nicholson and G. Butler et al., 1999. Possible role of trichothecene mycotoxins in virulence of Fusarium graminearum on maize. Plant Dis., 83: 954-960.
- Proctor, R.H., A.E. Desjardins, S.P. McCormick, R.D. Plattner, N.J. Alexander and D.W. Brown, 2002. Genetic analysis of the role of trichothecene and fumonisin mycotoxins in the virulence of Fusarium. Eur. J. Plant Pathol., 108: 691-698.
- Adams, G.C. and L.P. Hart, 1989. The role of deoxynivalenol and 15-acetyldeoxynivalenol in pathogenesis by Gibberella zeae, as elucidated through protoplast fusions between toxigenic and nontoxigenic strains. Phytopathology, 79: 404-408.
- Williams, L.D., A.E. Glenn, A.M. Zimeri, C.W. Bacon, M.A. Smith and R.T. Riley, 2007. Fumonisin disruption of ceramide biosynthesis in maize roots and the effects on plant development and Fusarium verticillioides-induced seedling disease. J. Agric. Food Chem., 55: 2937-2946.
- Arias, S.L., M.G. Theumer, V.S. Mary and H.R. Rubinstein, 2012. Fumonisins: Probable role as effectors in the complex interaction of susceptible and resistant maize hybrids and Fusarium verticillioides. J. Agric. Food Chem., 60: 5667-5675.
- van Asch, M.A.J., F.H.J. Rijkenberg and T.A. Coutinho, 1992. Phytotoxicity of fumonisin B1, moniliformin, and T-2 toxin to corn callus cultures. Phytopathology, 82: 1330-1332.
- Glenn, A.E., N.C. Zitomer, A.M. Zimeri, L.D. Williams, R.T. Riley and R.H. Proctor, 2008. Transformation-mediated complementation of a FUM gene cluster deletion in Fusarium verticillioides restores both fumonisin production and pathogenicity on maize seedlings. Mol. Plant-Microbe Interact., 21: 87-97.
- Desjardins, A.E., R.D. Plattner, T.C. Nelsen and J.F. Leslie, 1995. Genetic analysis of fumonisin production and virulence of Gibberella fujikuroi mating population A (Fusarium moniliforme) on maize (Zea mays) seedlings. Appl. Environ. Microbiol., 61: 79-86.
- Nel, B., C. Steinberg, N. Labuschagne and A. Viljoen, 2006. The potential of nonpathogenic Fusarium oxysporum and other biological control organisms for suppressing fusarium wilt of banana. Plant Pathol., 55: 217-223.
- Liu, L., J.W. Kloepper and S. Tuzun, 1995. Induction of systemic resistance in cucumber against fusarium wilt by plant growth-promoting rhizobacteria. Phytopathology, 85: 695-698.
- Gerrits, P.O. and R.W. Horobin, 1996. Glycol methacrylate embedding for light microscopy: Basic principles and trouble-shooting. J. Histotechnol., 19: 297-311.
- Beukes, I., L.J. Rose, G.S. Shephard, B.C. Flett and A. Viljoen, 2017. Mycotoxigenic Fusarium species associated with grain crops in South Africa-A review. S. Afr. J. Sci., 113.
- Leyva-Madrigal, K.Y., C.P. Larralde-Corona, M.A. Apodaca-Sánchez, F.R. Quiroz-Figueroa and P.A. Mexia-Bolaños et al., 2015. Fusarium species from the Fusarium fujikuroi species complex involved in mixed infections of maize in Northern Sinaloa, Mexico. J. Phytopathol., 163: 486-497.
- Elzein, A., A. Heller, B. Ndambi, M. de Mol, J. Kroschel and G. Cadisch, 2010. Cytological investigations on colonization of sorghum roots by the mycoherbicide Fusarium oxysporum f. sp. strigae and its implications for Striga control using a seed treatment delivery system. Biol. Control, 53: 249-257.
- Beckman, C.H., 1987. The Nature of Wilt Diseases of Plants. APS Press, Saint Paul, Minnesota, ISBN: 9780890540749, Pages: 175.
- Ruiz-Roldan, M.C., A. Di Pietro, M.D. Huartas-Gonzalez and M.I.G. Roncero, 1999. Two xylanase genes of the vascular wilt pathogen Fusarium oxysporum are differentially expressed during infection of tomato plants. Mol. Genet. Genomics, 261: 530-536.
- Jardine, D.J. and J.F. Leslie, 1999. Aggressiveness to mature maize plants of Fusarium strains differing in ability to produce fumonisin. Plant Dis., 83: 690-693.
- Leslie, J.F. and B.A. Summerell, 2006. The Fusarium Laboratory Manual. 1st Edn., Blackwell Publishing, Oxford, United Kingdom, ISBN: 9780813819198, Pages: 388.
How to Cite this paper?
APA-7 Style
Mwanja,
Y.P., Shutt,
V.M., Musa,
F., Ayika,
P.D. (2025). Infectivity of the Fusarium fujikuroi Species Complex (FFSC) on Maize (Zea mays Lam.) in Gauteng, South Africa. Research Journal of Botany, 20(1), 111-123. https://doi.org/10.3923/rjb.2025.111.123
ACS Style
Mwanja,
Y.P.; Shutt,
V.M.; Musa,
F.; Ayika,
P.D. Infectivity of the Fusarium fujikuroi Species Complex (FFSC) on Maize (Zea mays Lam.) in Gauteng, South Africa. Res. J. Bot 2025, 20, 111-123. https://doi.org/10.3923/rjb.2025.111.123
AMA Style
Mwanja
YP, Shutt
VM, Musa
F, Ayika
PD. Infectivity of the Fusarium fujikuroi Species Complex (FFSC) on Maize (Zea mays Lam.) in Gauteng, South Africa. Research Journal of Botany. 2025; 20(1): 111-123. https://doi.org/10.3923/rjb.2025.111.123
Chicago/Turabian Style
Mwanja, Yakubu, Pwajok, Vou Moses Shutt, Filibus Musa, and Philomena Daniel Ayika.
2025. "Infectivity of the Fusarium fujikuroi Species Complex (FFSC) on Maize (Zea mays Lam.) in Gauteng, South Africa" Research Journal of Botany 20, no. 1: 111-123. https://doi.org/10.3923/rjb.2025.111.123

This work is licensed under a Creative Commons Attribution 4.0 International License.