Effect of Trichoderma Species on Seed Borne Fungi of Pearl Millet (Pennisetum glaucum L.R. Br) Using Dual Culture Technique
-
Liamngee Kator


Department of Biological Sciences, Benue State University, Makurdi, Nigeria
Fayinminu Akintade OjoDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
Anyebe Benard OcheiduduDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
Akula Helen NgodooDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
Yavaku Joy DoowueseDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
Achagh Terpase LeviDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
Emmanuel Dorcas EneDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
Ikwuba Favour OnahiDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
Atsor Linda NguemoDepartment of Biological Sciences, Benue State University, Makurdi, Nigeria
| Received 23 Nov, 2024 |
Accepted 13 Jan, 2025 |
Published 15 Jan, 2025 |
Background and Objective: Millets are susceptible to fungi diseases resulting in high postharvest losses. To minimize these losses, there is a need for effective and eco-friendly control measures towards these pathogens. Thus, the study was carried out to investigate the effect of Trichodermaspecies on seed-borne fungi of pearl millet (Pennisetum glaucum)in Makurdi. Materials and Methods: Pearl millet seeds were collected from three markets in the Makurdi metropolis namely, Wurukum, Railway, and High-Level Markets, and taken to the Botany Laboratory of the Benue State University for the isolation of fungi. Soil samples from millet plantations in the Apir area of Makurdi were collected in polythene bags and taken to the Laboratory for the isolation of Trichodermaspecies. The standard blotter method was employed for the detection and isolation of seed-borne fungi in pearl millet and the serial dilution technique was used for the isolation of Trichodermafrom soil samples. The effect of Trichoderma koningiion the radial growth of isolated fungi was carried out using a dual culture technique. Data collected from the study were analyzed using Chi-square and t-test. Means were separated using Duncan’s Multiple Range Test (DMRT) at a 5% level of significance. Results: The study identified four seed-borne fungi in pearl millet: Aspergillus flavus, Aspergillus niger, Rhizopus stolonifer, and Fusarium sp. A significant location-based variation in fungal occurrence was observed, with Aspergillus flavus being the most prevalent. Seed germination showed no significant difference across locations. Fungal growth was significantly reduced when paired with Trichoderma koningii, with Rhizopus stolonifer showing the highest reduction in growth compared to other fungi. Conclusion: Trichoderma koningii proved to be an effective bio-control agent as it inhibited the radial growth of fungi isolates and can be employed in plant disease management. Therefore, T. koningii should be employed for seed treatment and industries should be created by the Government and relevant stakeholders where Trichodermaisolates can be formulated and produced on a commercial scale for farmers.
| Copyright © 2025 Kator et al. This is an open-access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. |
INTRODUCTION
Pearl millet (Pennisetum glaucum L. R. Br.), also known as bajra in Hindu or ‘yadi’ in the Marghi language of North Eastern Nigeria, is a cereal crop of paramount importance, exhibiting considerable potential among millet varieties1. Characterized by its robust growth habit, pearl millet is a dual-purpose crop, offering both grain and fodder, with exceptional yielding potentials. As a staple food, it plays a vital role in supporting millions of people in arid and semi-arid regions worldwide2. According to the Food and Agriculture Organization (FAO), pearl millet is the sixth most important cereal crop globally, with significant annual cultivation in Africa and the Indian Sub-Continent3. India is the largest producer of pearl millet in Asia, with an average productivity of 930 kg/ha. Nigeria ranks as the fifth largest producer of millet worldwide, with an annual production of 1.5 million tonnes as of 20164.
Pearl millet is a staple food in many Nigerian households, particularly among low-income families in Northern Nigeria, with millions of tonnes consumed annually3. The crop is also utilized in the production of various traditional food products, including “masa” (a fried cake) and “tuwo” (a thick binding paste). Furthermore, pearl millet flour is used to prepare flatbreads, leveraging its gluten-free properties.
Nutritionally, pearl millet is a valuable source of protein (18%), vitamins B, particularly niacin, B6, and folic acid. Its importance extends across the Sahel Region, serving as a primary staple in Northern Nigeria, Niger, Mali, and Burkina Faso. In some regions, pearl millet is processed into a beverage called “fura” (Hausa) or “tukura” (Marghi), which involves grinding the grain into flour, rolling it into balls, parboiling, and mixing it with fermented milk to create a watery paste1.
Millet crops are susceptible to various diseases, which can manifest severely under diverse climatic conditions, resulting in substantial economic losses5. While some diseases occur sporadically in specific climates with minimal impact, others are more prevalent and detrimental. Notably, fungal diseases predominate over bacterial and viral diseases in millets5. Key fungal diseases affecting millets include grain mold, ergot, smut, anthracnose, downy mildew, blast, rust, charcoal rot, foot rot, banded sheath blight, and sheath rot. These diseases infect various plant parts, such as roots, stems, leaves, peduncles, and grains, ultimately compromising yield and quality5. Furthermore, these fungal diseases not only cause significant qualitative and quantitative yield reductions but also produce mycotoxins, posing serious health risks to humans and animals that consume contaminated grains6.
The various methods used for managing disease include the use of resistant varieties, cultural practices, chemical control, and biological control. Breeding for disease-resistant varieties has been long used for managing diseases7. The evolution of pathogens into newer, more aggressive biotypes poses a significant challenge to disease management in crops7. This phenomenon can lead to the breakdown of resistance in previously resistant varieties, rendering them susceptible to infection.
Traditionally, chemical pesticides, fungicides, and microbicides, such as fthalide, edifenphos, iprobenfos, tricyclazole, isoprothiolane, probenazole, pyroquilon, felimzone (meferimzone), and diclocymet, have been employed to manage diseases in crops. While these chemicals are effective, their use is often accompanied by significant economic and environmental costs, as well as potential human health risks8. The use of alternate methods instead of seed-treating chemicals is of great concern to save our environment. Seed treatment with different plant extracts has been shown effective in controlling seed-borne fungal pathogen9. However, this study seeks to evaluate a biological control method where antagonistic organisms will be used to control fungal pathogens of millet in vitro. Therefore, the effect of Trichoderma species on seed-borne fungi of pearl millet in Makurdi using the dual culture technique was investigated.
MATERIALS AND METHODS
Study area: The study was carried out at the Plant Science and Biotechnology Laboratory of the Benue State University between November, 2023 to May, 2024.
Collection of pearl millet samples: Pearl millet was collected from three major markets in Makurdi namely; Wurukum, Railway, and High-level markets. The millet seeds were collected from three different sale points in each market. They were packaged in polythene bags, labeled properly, and taken to the Botany Laboratory of Benue State University for isolation of the seed-borne fungi.
Preparation of culture media: Potato dextrose agar (PDA) media was used for the isolation of the fungal pathogens for further studies. The media was prepared according to the manufacturer’s recommended procedures where 39.6 g of powdered PDA medium were dissolved in 1000 mL of sterile distilled water and stirred vigorously to homogenize. The content of the flask was heated on a heating mantle until the solution became clear and all the PDA powder dissolved. After heating, the mouth of the flask was covered with cotton wool and autoclaved at 121°C for 15 min at 760 mmHg. The sterile medium was allowed to cool to a temperature at which it could be held with hands and two to three drops of streptomycin sulphate were added to inhibit bacterial growth. The medium was dispensed into the Petri dishes and allowed to solidify for further studies.
Detection of fungi on millet seeds: The detection of seed-borne fungi was done using the blotter method10. Three layers of Whatman filter papers were soaked in sterile distilled water and placed at the bottom of 9 cm diameter Petri dishes. The seeds were surface sterilized in 5% sodium hypochlorite for 30 sec to 1 min and rinsed in three successive changes of sterile distilled water11. Fifteen sterilized seeds were placed on the moist filter paper in the Petri dishes and incubated at ambient temperature for 5-7 days. After 5-7 days of incubation, the seeds were examined for fungal growth and germination of seeds. The data collected include.
Percentage occurrence of fungi: Plates were observed for growth and the occurrence of fungi was determined by counting the number of fungi per market divided by the total number of fungi and expressed as a percentage12:
Percentage occurrence of specific fungi: Seeds were observed for growth and the occurrence of specific fungi was determined by counting the number of times each fungus occurred divided by the total number of fungi and expressed as a percentage using the formula12:
Percentage of seed germination: The percentage of seed germination was calculated by counting the number of seeds with seed leaves divided by the total number of seeds per plate12:
Isolation and sub-culturing of fungi from millet seeds: On the appearance of fungi, a small quantity of each fungi colony was picked with the aid of an inoculation needle and inoculated on prepared potato dextrose agar. The Petri plates were incubated at ambient conditions of light and temperature for 5-7 days and observed daily for fungal growth. After 5-7 days, sub-culturing was done to obtain the pure culture
of the isolates. To subculture, a sterilized inoculation needle was used to pick a small quantity of the fungal growth on the old culture and transfer it to the center of a freshly prepared PDA in another Petri dish. Sub-culturing was done repeatedly until pure cultures of each fungal organism were obtained as reported by Kator et al.12.
Identification of fungi: The identification of fungi was done by observing the color, nature of growth, and growth rate of the fungi macroscopically in Petri plates. Microscopic identification was done by staining a glass slide with a drop of Lactophenol in cotton blue and with the aid of an inoculation needle, a small quantity of the fungal colony was placed on the stained-glass slide. This was covered with a cover slip and viewed under the 40× objective lens of the light microscope. The observed characteristic of the fungi was compared with standard text for identification12,13.
Collection of soil samples: Soil samples were collected from millet plantations in the Apir Area of Makurdi, Benue State. The soil samples within the area of the millet plants were collected in polyethylene bags at a depth of 2-3 cm from the rhizosphere of the plants at three different points on the field and pulled together14. These were taken to the Botany Laboratory of the Benue State University for isolation of Trichoderma species.
Isolation of Trichoderma species from soil samples: The serial dilution method of Liamngee et al.14 was employed for this study. For isolation of Trichoderma species from the soil samples, a serial dilution technique was employed and a 104 dilution of the soil sample was prepared. In this method, a stock suspension was prepared by adding 1 g of the soil samples to 9 mL of sterile distilled water in a sterilized glass tube. One milliliter was further pipetted from the first dilution and introduced into another labelled test tube to obtain 10–1 dilution. This was done repeatedly to obtain up to 10–4 dilution. A 10–4 dilution of the sample was prepared and 1 mL was dispersed in a 9 cm diameter Petri dish. Approximately 15-20 mL of sterilized, molten potato dextrose agar (PDA) was then added to the dish. The agar and inoculum were gently swirled to ensure uniform distribution, and the mixture was allowed to solidify. The culture plates were incubated at room temperature for 7 days. Daily observations were made, and each colony that emerged was considered a single colony-forming unit (CFU). Pure cultures of fungal colonies were obtained through subculturing, following the procedure outlined in section 2.4.4.
Identification of Trichoderma isolates: Identification of Trichoderma species was conducted through visual observation on Petri dishes and micro-morphological studies using slide culture14. Visual observations on PDA-grown isolates focused on growth rates, changes in medium color, and colony morphology, which are considered taxonomically informative characteristics for Trichoderma. The micro-morphological examination involved observing the morphology of conidiophores and conidia, as well as other relevant characteristics, using electronic documentation on the genus Trichoderma14.
In vitro antagonistic activity of Trichoderma species: The in vitro antagonistic effects of Trichoderma species against test fungi (isolated from millet seeds) were evaluated using the dual culture technique on potato dextrose agar (PDA) medium14. Mycelial discs (5 mm) of Trichoderma species and test pathogens were placed simultaneously, 1 cm from the edge of each Petri dish, in opposite directions. The experiment was conducted in triplicate, with a completely randomized design. Control plates received only the mycelial disc of each test fungus. Plates were incubated at ambient laboratory temperature. The inhibition of test fungi was calculated using the following formula14:
Where:
| R1 | = | Mycelia growth of the pathogen without Trichoderma (control) | |
| R2 | = | Mycelia growth of the pathogen in the presence of Trichoderma |
Data analysis: Data collected from the study were analyzed using analysis of variance and t-test. Means were separated using Duncan’s Multiple Range Test at a 5% level of significance.
RESULTS
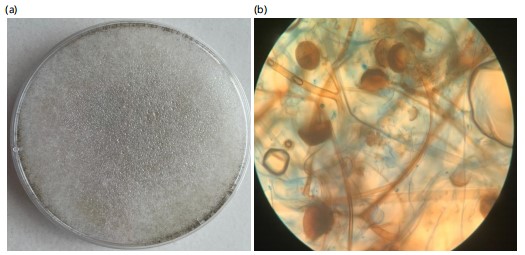
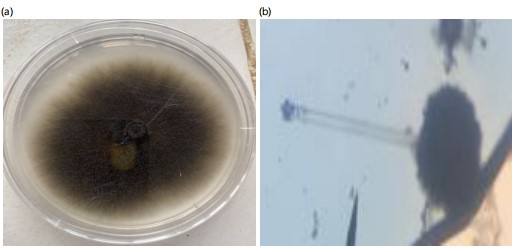
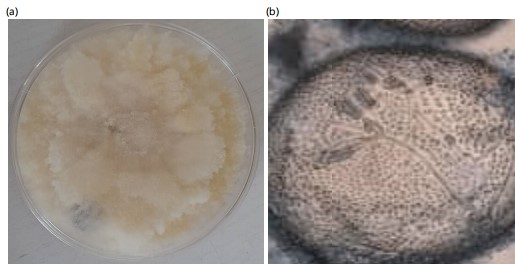
The study revealed that four fungi were isolated from pearl millets in Makurdi while Trichoderma koningii was isolated from the soil sample. Their macroscopic and microscopic features are presented in Fig. 1-5. The colony of A. flavus on PDA had a greenish coloration (Fig. 1a). When viewed under the microscope, A. flavus had bluish-green conidia with the conidiophore’s hyaline in nature (Fig. 1b). The colonies of R. stolonifer were fast growing with cottony like appearance on PDA (Fig. 2a). The sporangiospores were hyaline, smooth-walled and branched forming large terminal globose sporangia (Fig. 2b). Colony of Aspergillus niger on PDA was black (Fig. 3a) and the conidiophores were hyaline, inflated at the apex forming globose conidia vesicles when viewed under the microscope (Fig. 3b). The colony color of Fusarium sp., was pinkish white a cottony like appearance on PDA (Fig. 4a). The conidia had a cylindrical shape with a septum, single cell, hyaline and a smooth round end when viewed under the light microscope (Fig. 4b). Trichoderma koningii on PDA had a rapid growth and produced yellowish-white mycelia with rough surface (Fig. 5a). When viewed under the microscope, the conidia were ellipsoidal, hung at the apex of a long conidiophore, and branched at alternate angles Fig. 5b.

|
|
|

|
|
Occurrence of seed borne fungi on pearl millet based on location: The occurrence of seed-borne fungi on pearl millet is shown in Table 1. There was a significant relationship in fungi occurrence based on location (χ2 = 14.262, df = 6, p = 0.027). Aspergillus flavus had significantly higher occurrence with 16 (47.06%), followed by Aspergillus niger 9 (26.47%), Fusarium sp., 7 (20.59%) and the least was Rhizopus stolonifer 2 (5.88%). Pearl millet seeds obtained from High Level had significantly higher fungi occurrence with 13 (38.24%) followed by Railway 11 (32.35%) and the least was Wurukum 10 (29.41%).
Percentage seed germination of pearl millet in Makurdi: The percentage of seed germination of pearl millet is presented in Table 2. Out of the 135 seeds that were plated, 103 seeds representing 76.30% notably germinated. There was no significant seed germination in pearl millet collected from Wurukum, Railway, and high-level markets (χ² = 3.522, df = 2, p = 0.172). Pearl millet seeds obtained from high levels had the highest seed germination with 37 (27.41%) followed by Wurukum at 36 (26.67%) and the least was from Railway with 30 (22.22%).
Effect of Trichoderma koningii on radial growth of Rhizopus stolonifer isolated from pearl millet: The effect of Trichoderma koningii on radial growth of Rhizopus stolonifer is presented in Table 3. There was no significant difference in the radial growth of Rhizopus stolonifer in the control and radial growth of Rhizopus stolonifer paired with Trichoderma koningii after 48 hrs of observation. However, there was a significantly higher radial growth in the control (2.13±0.15, 2.83±0.21 and 3.57±0.42) cm at 72, 96, and 120 hrs, respectively compared with radial growth of Rhizopus stolonifer paired with Trichoderma koningii (1.50±0.10, 1.78±0.10 and 2.15±0.30) at 72 to 120 hrs, respectively.
Effect of Trichoderma koningii on radial growth of Aspergillus flavus Isolated from pearl millet: The effect of Trichoderma koningii on radial growth of Aspergillus flavus is presented in Table 4. Radial growth of Aspergillus flavus paired with Trichoderma koningii was significantly lower at 48 to 120 hrs (0.67±0.13, 0.88±0.38, 0.93±0.38 and 1.17±0.21), respectively compared with the control (1.70±0.43, 2.30±0.52, 2.84±0.80 and 3.47±1.15) at 48 to 120 hrs.
| Table 1: | Occurrence of seed borne fungi of pearl millet in Makurdi Metropolis | |||
| Location | ||||
| Fungi isolate | Wurukum | Railway | High level | Total |
| Rhizopus stolonifera | 1 | 1 | 0 | 2 |
| Aspergillus flavus | 7 | 6 | 3 | 16 |
| Fusarium sp. | 0 | 4 | 3 | 7 |
| Aspergillus niger | 2 | 0 | 7 | 9 |
| Total (%) | 10(29.41) | 11(32.35) | 13(38.24) | 34 |
| χ2 = 14.262, df = 6 and p = 0.027 | ||||
| Table 2: | Percentage seed germination of pearl millet in Makurdi | |||
| Location | Number of seeds germinated (n=135) | Germination (%) |
| Wurukum | 36 | 26.67 |
| Railway | 30 | 22.22 |
| High level | 37 | 27.41 |
| Total | 103 | 76.3 |
| Table 3: | Effect of Trichoderma koningii on radial growth of Rhizopus stolonifer isolated from pearl millet | |||
| Radial growth (cm) | ||||
| Treatment | 48 hrs | 72 hrs | 96 hrs | 120 hrs |
| R. stolonifer+T. koningii | 1.07±0.12 | 1.50±0.10 | 1.78±0.10 | 2.15±0.30 |
| Control | 1.67±0.40 | 2.13±0.15 | 2.83±0.21 | 3.57±0.42 |
| T-value | 2.472 | 6.008 | 7.814 | 4.759 |
| p-value | 0.069 | 0.004 | 0.001 | 0.009 |
| Values are Mean±Standard Deviation in triplicates | ||||
| Table 4: | Effect of Trichoderma koningii on Aspergillus flavus isolated from pearl millet | |||
| Radial growth (cm) | ||||
| Treatment | 48 hrs | 72 hrs | 96 hrs | 120 hrs |
| A. flavus+T. koningii | 0.67±0.31 | 0.88±0.38 | 0.93±0.38 | 1.17±0.21 |
| Control | 1.70±0.43 | 2.30±0.52 | 2.84±0.80 | 3.47±1.15 |
| T-value | 3.369 | 3.784 | 3.708 | 3.408 |
| p-value | 0.028 | 0.019 | 0.021 | 0.027 |
| Values are Mean±Standard Deviation in triplicates | ||||
| Table 5: | Effect of Trichoderma koningii on Fusarium sp. isolated from pearl millet | |||
| Radial growth (cm) | ||||
| Treatment | 48 hrs | 72 hrs | 96 hrs | 120 hrs |
| Fusarium sp.+T. koningii | 0.15±0.05 | 0.23±0.11 | 0.47±0.25 | 0.68±0.43 |
| Control | 2.93±0.90 | 3.47±0.85 | 3.83±0.90 | 4.17±1.26 |
| T-value | 5.337 | 6.539 | 6.22 | 4.511 |
| p-value | 0.006 | 0.003 | 0.003 | 0.011 |
| Values are Mean±Standard Deviation in Triplicates | ||||
| Table 6: | Effect of Trichoderma koningii on Aspergillus niger isolated from pearl millet | |||
| Radial growth (cm) | ||||
| Treatment | 48 hrs | 72 hrs | 96 hrs | 120 hrs |
| A. niger+T. koningii | 0.83±0.29 | 1.17±0.35 | 1.47±0.42 | 1.76±0.25 |
| Control | 2.02±0.08 | 5.00±0.40 | 5.10±0.53 | 6.73±1.55 |
| T-value | 6.947 | 12.293 | 9.347 | 5.472 |
| p-value | 0.002 | 0.000 | 0.001 | 0.005 |
| Values are Mean±Standard Deviation in triplicates | ||||
| Table 7: | Comparative effect of Trichoderma koningii on radial growth of fungi species isolated from pearl millet | |||
| Radial growth (cm) | ||||
| Treatment | 48 hrs | 72 hrs | 96 hrs | 120 hrs |
| R. stolonifer+T. koningii | 1.07±0.12a | 1.50±0.10d | 1.78±0.10ac | 2.15±0.30g |
| A. flavus+T. koningii | 0.67±0.31b | 0.88±0.38e | 0.93±0.38c | 1.17±0.21i |
| Fusarium sp.+T. koningii | 0.15±0.05d | 0.23±0.11def | 0.47±0.25ab | 0.68±0.43gh |
| A. niger+T. koningii | 0.83±0.29c | 1.17±0.35f | 1.47±0.42b | 1.76±0.25h |
| Values are Mean±Standard Deviation in triplicates, mean values with similar alphabets are not significant, otherwise they are | ||||
Effect of Trichoderma koningii on radial growth of Fusarium sp., isolated from pearl millet: The effect of Trichoderma koningii on radial growth of Fusarium sp., is presented in Table 5. Radial growth of Fusarium sp., paired with Trichoderma koningii was significantly lower at 48 to 120 hrs (0.15±0.05, 0.23±0.11, 0.47±0.25 and 0.068±0.43), respectively compared with the control (2.93±0.90, 3.47±0.85, 3.84±0.90 and 4.17±1.26) at 48 to 120 hrs.
Effect of Trichoderma koningii on radial growth of Aspergillus niger isolated from pearl millet: The effect of Trichoderma koningii on radial growth of Aspergillus niger is presented in Table 6. Radial growth of Aspergillus niger paired with Trichoderma koningii was significantly lower at 48 to 120 hrs (0.83±0.29, 1.17±0.35, 1.47±0.42 and 1.76±0.25), respectively compared with the control (2.02±0.08, 5.00±0.40, 5.10±0.53 and 6.73±1.55) at 48 to 120 hrs.
Comparative effect of Trichoderma koningii on radial growth of fungi species isolated from pearl millet: The comparative effect of Trichoderma koningii on radial growth of fungi species isolated from pearl millet is presented in Table 7. Radial growth of Rhizopus stolonifer paired with Trichoderma koningii was significantly higher at 48 to 120 hrs (1.07±0.129, 1.50±0.10, 1.78±0.10 and 2.15±0.30) compared with radial growth of Aspergillus niger (0.83±0.29, 1.17±0.35, 1.47±0.42 and 1.76±0.25), Aspergillus flavus (0.67±0.31, 0.88±0.38, 0.93±0.38 and 1.17±0.21), and Fusarium sp. (0.15±0.05, 0.23±0.11, 0.47±0.25 and 0.68±0.43), respectively.
DISCUSSION
The seed-borne fungi isolated from pearl millet in this study were; Aspergillus flavus, Aspergillus niger, Rhizopus stolonifer, and Fusarium sp. A study by Hussain et al.15 isolated similar fungal organisms which include; Alternaria alternata, Aspergillus alba, A. flavus, A. niger, Bipolaris spp., C. lunata, Drechslera spp., Fusarium semitectum, F. moniliforme, Helminthosporium spp., Penicillium spp., and Rhizopus spp., from different varieties of millets in Pakistan. A similar study by Malik et al.3 in Kebbi State Nigeria reported fungi species of Aspergillus fumigatus, Aspergillus niger, Grapium sp., Microsporum sp., Trichophyton sp., as seed borne fungi from millets. Also in Kano State Nigeria, Fayinminu et al.6 implicated fungi species of A. flavus, A. fumigatus, A. niger, A. oryzae, A. Candida, and A. Parasiticus as seed-borne fungi from millet. Seed-borne pathogens have been involved in seed rots during germination and seedling mortality leading to poor crop stand. Seed-borne pathogens, whether present externally or internally, can have devastating effects on plant growth and productivity. These pathogens can cause a range of symptoms, including seed abortion, seed rot, seed necrosis, and reduced or eliminated germination capacity. Additionally, infected seeds can give rise to damaged seedlings, which can lead to the development of disease at later stages of plant growth through systemic or local infection16. The presence of fungal pathogens in millet seeds suggests contamination occurred during field harvesting. Previous studies have reported the association of field and storage fungal pathogens with millet seeds. Fungal pathogens, including Aspergillus niger, Penicillium sp., Fusarium sp., Rhizopus sp., and Helminthosporium sp., have been isolated from rotted seeds and abnormal seedlings, and have been implicated in seed decay and damping-off of millet seedlings17.
The presence of storage fungal genera, such as Aspergillus and Penicillium, on seeds indicates that contamination occurred during storage. The detection of A. niger and A. flavus on seeds, abnormal seedlings, and rotted seeds confirms that Aspergillus species, although often saprophytic, can cause reduced germination in seeds18. Furthermore, A. flavus produces toxic metabolites that can inhibit shoot and root elongation18. Aspergillus niger is a damaging storage fungus that can deteriorate seed quality and reduce seed germination. Moreover, molds associated with pearl millet seeds have been reported to be pathogenic to humans, producing toxins that may cause respiratory diseases15.
The study reported that pearl millet seeds obtained from high-level markets had the highest number of fungi compared to seeds obtained from Wurukum and Railway markets. The disparity observed in the occurrence of fungi in the different locations could be a result of the environmental conditions of the markets, the sanitary condition, the method of storage used, and the mode of display for sale in the market. These pathogens associated with millet seeds must have been present right from the field and after harvest, transferred to storage houses and marketplaces19. Also, the study revealed that millet seeds collected from High levels had the highest percentage of germination compared to seeds obtained from other markets. The findings from this study conforms with other research findings of Hussain et al.15 and Makun et al.17 in which they reported different percentages seed germination of cereals obtained from different locations. The variation in percentage seed germination within the locations as observed in this study might be a result of the source of collection of the millet seeds, the storage method employed, and also the possibility of the seed treatment technique used.
In terms of individual fungi occurrence, Aspergillus flavus was implicated in this study to be the dominant fungi isolated from millet seeds obtained across the locations. This result was contrary to the previous findings3,6,15 in which they reported fungi species of Trichophyton sp., Alternaria alternata, and Aspergillus niger as the dominant seed-borne fungi pathogens of millet, respectively. The percentage occurrence of A. flavus (26.47%) recorded in this study was high compared to 14.00 and 11.67% reported by Fayinminu et al.6 and Hussain et al.15, respectively. The variability in the frequency of occurrence of these fungi may be attributed to differences in inoculum density in the area or prevailing environmental conditions that favor fungal growth20. These fungi likely colonized the grains at various stages, including during production in the field, transportation or storage.
The effect of Trichoderma koningii on radial growth of the isolated fungi from pearl millet in this study revealed that the different fungi gave different radial growth but T. koningii was able to inhibit their radial growth compared to untreated fungi. The use of T. koningii in the control of fungi radial growth in the study is a diversification away from other previous methods of control such as the use of plant extract reported by several researchers. In a similar study carried out by Khalili21, it was reported that Trichoderma harzianum and Trichoderma virens were able to significantly inhibit the mycelial growth of Bipolaris oryzae. He et al.22 stated that disease reduction by biological control is possible by reducing pathogen inoculum (decreased development and release of viable spores, decreased survival, and reduced spread), reducing pathogen infection of the host, and reducing the severity of the pathogen attack. The ability of T. koningii to inhibit the growth of the isolated fungi could be attributed to its ability to grow faster than the other organisms implying the need for more nutrient utilization and space than them. This reason is supported by Khalili et al.21, who explained that inhibition of pathogen growth is due to the production of amylase by Trichoderma spp., which is partially responsible for the rapid growth of antagonists in potato dextrose agar medium. Liamngee et al.14 added that in addition to amylase, it has been reported that Trichoderma species also produce extracellular cellulose and pectinase enzymes that are capable of hydrolyzing the cell walls of other fungi.
CONCLUSION
The study has shown that A. niger, A. flavus, R. stolonifer, and Fusarium sp., were fungi isolated from seeds of pearl millet in Makurdi. The distribution of these fungi in millet seeds varied across locations with seeds obtained from High levels having the highest fungi occurrence. The dominant species was A. flavus in pearl millet seeds collected across the locations. Trichoderma koningii proved to be an effective biocontrol agent as it inhibited the radial growth of the fungal isolates. Based on the study’s findings, it is recommended that additional research be conducted on seed-borne fungi in pearl millet, an area with limited investigation. Further studies should also focus on isolating other Trichoderma species from the rhizosphere of pearl millet plantations for documentation. Additionally, Trichoderma koningii should be formulated into powders and produced on a commercial scale for use by farmers.
SIGNIFICANCE STATEMENT
High-quality seed is important for increased crop production and the proper establishment of a sound seed industry in the country. Trichoderma koningii used in the study significantly inhibited fungi growth in vitro. Therefore, the study’s outcome proves relevant to the government, farmers, researchers, and consumers. For researchers, it will provide the basis for more research on other biocontrol agents and how they can be formulated for farmers. For consumers, it will provide them with safe and healthy produce for consumption. For policymakers, it will enable them to enact laws favoring the use of biocontrol agents in plant disease control. The government could also create industries where these biocontrol agents can be formulated and produced in large quantities thereby creating employment.
REFERENCES
- Izge, A.U. and I.M. Song, 2013. Pearl millet breeding and production in Nigeria: Problems and prospects. J. Environ. Issues Agric. Dev. Countries, 5: 25-33.
- Chopra, V.L., 2001. Breeding Field Crops. Oxford Ibh, Oxford and IBH Publishing House, New Delhi, India, ISBN-13: 9788120414358, Pages: 580.
- Malik, V.K., P. Sangwan, M. Singh, P. Kumari and N. Shoeran et al., 2024. Stem rot of pearl millet prevalence, symptomatology, disease cycle, disease rating scale and pathogen characterization in pearl Millet-Klebsiella pathosystem. Plant Pathol. J., 40: 48-58.
- Abah, D., E.O. Oraka and O.C. Ochoche, 2021. Trend analysis of millet production and agricultural output in Nigeria (1981-2018). Int. Res. J. Innovations Eng. Technol., 5: 5-10.
- Poonacha, T.T., C.H.S. Bhavana, Farooqkhan, G.V. Ramesh and N. Gavayi et al., 2023. Blast Disease of Millets: Present Status and Future Perspectives. In: Millets-Rediscover Ancient Grains, Yadav, L. and U. Upasana (Eds.), IntechOpen, London, United Kingdom, ISBN: 978-1-83768-701-5.
- Fayinminu, A.O., T.T. Ati, B.L. Agbatar, S.J. San and J.J. Orbunde, 2020. Incidence of Aspergillus species in maize seeds in Konshisha Local Government of Benue State and their control using neem leaf extract. Biol. Life Sci., 53.
- Paul, P., M. Kabilan, V. Duraipandiyan, V.P. Rani, D.A. Prabhu and P. Oviya, 2023. A review on disease management in millets. Int. J. Sci. Res. Sci. Technol., 10: 315-328.
- Limtong, S., P. Into and P. Attarat, 2020. Biocontrol of rice seedling rot disease caused by Curvularia lunata and Helminthosporium oryzae by epiphytic yeasts from plant leaves. Microorganisms, 8.
- Mohana, D.C., P. Prasad, V. Vijaykumar and K.A. Raveesha, 2011. Plant extract effect on seed-borne pathogenic fungi from seeds of paddy grown in Southern India. J. Plant Prot. Res., 51: 101-106.
- Akonda, M.M.R., M. Yasmin and I. Hossain, 2016. Incidence of seedborne mycoflora and their effects on germination of maize seeds. Int. J. Agron. Agric. Res., 8: 87-92.
- Ekefan, E.J., A.O. Nwankiti and V.I. Gwa, 2018. Comparative assessment of antimicrobial potency of some selected plant extracts against seed borne pathogens of germinating yam setts. J. Plant Pathol. Microbiol., 9.
- Kator, L., A.T. Ogo-Oluwa and A.B. Kemi, 2016. Isolation and identification of seed borne fungi of common bean (Phaseolus vulgaris L.) from selected markets in Makurdi. Int. J. Appl. Agric. Sci., 2: 75-78.
- Barnett, H.L. and B.B. Hunter, 1972. Illustrated Genera of Imperfect Fungi. 3rd Edn., Burgess Publishing Company, Minneapolis, Minnesota, ISBN: 9780808702665, Pages: 241.
- Liamngee, K., Y. Awua, A.O. Fayinminu and S.J. Anshi, 2023. Identification of fungi colonizing the rhizopshere of tomato (Solanum lycopersicum) plants and their pathogenicity on healthy tomato fruits. Niger. Ann. Pure Appl. Sci., 6: 186-194.
- Hussain, A., S.A. Anwar, G.M. Sahi, Q. Abbas and Imran, 2009. Seed borne fungal pathogens associated with pearl millet (Pennisetum typhoides) and their impact on seed germination. Pak. J. Phytopathol., 21: 55-60.
- Khanzada, K.A., M.A. Rajput, G.S. Shah, A.M. Lodhi and F. Mehboob, 2002. Effect of seed dressing fungicides for the control of seedborne mycoflora of wheat. Asian J. Plant Sci., 1: 441-444.
- Makun, H.A., T.A. Gbodi, A.S. Tijani, A. Abai and G.U. Kadiri, 2007. Toxicologic screening of fungi isolated from millet (Pennisetum spp) during the rainy and dry harmattan seasons in Niger State, Nigeria. Afr. J. Biotechnol., 6: 34-40.
- Gebeyaw, M., 2020. Review on: Impact of seed-borne pathogens on seed quality. Am. J. Plant Biol., 5: 77-81.
- Lamichhane, K., A.R. Khadka, R.P. Mainali and A. Karkee, 2023. Seed borne fungal pathogens associated with finger millet accessions. J. Plant Prot. Soc., 8: 60-75.
- Mahmoud, M.A., M.R. Al-Othman and A.R.M. Abd El-Aziz, 2013. Mycotoxigenic fungi contaminating corn and sorghum grains in Saudi Arabia. Pak. J. Bot., 45: 1831-1839.
- Khalili, E., M. Sadravi, S. Naeimi and V. Khosravi, 2012. Biological control of rice brown spot with native isolates of three Trichoderma species. Braz. J. Microbiol., 43: 297-305.
- He, D.C., M.H. He, D.M. Amalin, W. Liu, D.G. Alvindia and J. Zhan, 2021. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens, 10.
How to Cite this paper?
APA-7 Style
Kator,
L., Ojo,
F.A., Ocheidudu,
A.B., Ngodoo,
A.H., Doowuese,
Y.J., Levi,
A.T., Ene,
E.D., Onahi,
I.F., Nguemo,
A.L. (2025). Effect of Trichoderma Species on Seed Borne Fungi of Pearl Millet (Pennisetum glaucum L.R. Br) Using Dual Culture Technique. Research Journal of Botany, 20(1), 55-65. https://doi.org/10.3923/rjb.2025.55.65
ACS Style
Kator,
L.; Ojo,
F.A.; Ocheidudu,
A.B.; Ngodoo,
A.H.; Doowuese,
Y.J.; Levi,
A.T.; Ene,
E.D.; Onahi,
I.F.; Nguemo,
A.L. Effect of Trichoderma Species on Seed Borne Fungi of Pearl Millet (Pennisetum glaucum L.R. Br) Using Dual Culture Technique. Res. J. Bot 2025, 20, 55-65. https://doi.org/10.3923/rjb.2025.55.65
AMA Style
Kator
L, Ojo
FA, Ocheidudu
AB, Ngodoo
AH, Doowuese
YJ, Levi
AT, Ene
ED, Onahi
IF, Nguemo
AL. Effect of Trichoderma Species on Seed Borne Fungi of Pearl Millet (Pennisetum glaucum L.R. Br) Using Dual Culture Technique. Research Journal of Botany. 2025; 20(1): 55-65. https://doi.org/10.3923/rjb.2025.55.65
Chicago/Turabian Style
Kator, Liamngee, Fayinminu Akintade Ojo, Anyebe Benard Ocheidudu, Akula Helen Ngodoo, Yavaku Joy Doowuese, Achagh Terpase Levi, Emmanuel Dorcas Ene, Ikwuba Favour Onahi, and Atsor Linda Nguemo.
2025. "Effect of Trichoderma Species on Seed Borne Fungi of Pearl Millet (Pennisetum glaucum L.R. Br) Using Dual Culture Technique" Research Journal of Botany 20, no. 1: 55-65. https://doi.org/10.3923/rjb.2025.55.65

This work is licensed under a Creative Commons Attribution 4.0 International License.